КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Г. Электромиографические индикаторы динамичности тормозного процесса 3 страница
|
|
|
|
В соответствии с общим принципом сенсорной неспецифичности ретикулярных активаций последние могут быть вызваны раздражителями любой сенсорной модальности. В наших опытах мы пользовались звуковыми и световыми раздражителями. Оба эти вида стимулов могут вызвать реакцию активации (блокады, подавления альфа-ритма), однако между ними имеется и существенное различие, заключающееся в том, что реакция на звук достаточно быстро подвергается угашению, в то время как при реакции на свет угашению подвергается лишь следовая, десинхронизация, а реакция во время действия светового раздражителя при достаточной его интенсивности может сохраняться неопределенно долго. Это последнее обстоятельство несколько выделяет «световую» активацию из класса ориентировочных реакций, что молча признается большинством авторов, однако не ставит ее особняком настолько, чтобы вовсе исключить ее из этой категории. Действительно, как мы сейчас увидим, разница между ЭЭГ ориентировочными реакциями на звук и на свет имеет скорее количественный, чем качественный характер.
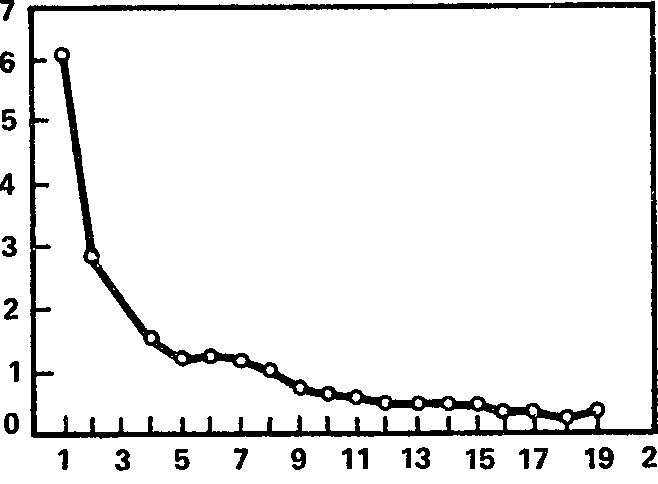
Рис. 15. Динамика угашения ориентировочной электрокорковой реакции на звук (в среднем для всей выборки). Ось абсцисс – порядковый номер предъявления звукового раздражителя; ось ординат – длительность реакции десинхронизации (в с).
Общее представление о динамике ориентировочной реакции на звук можно получить из рис. 15, на котором изображена средняя для выборки из 47 испытуемых длительность блокады альфа-ритма в ответ на последовательные предъявления звукового раздражителя (характеристики раздражителей указаны в гл. 3). Видно, что на первое предъявление звука десинхронизация длится в среднем более 6 с, т. е. к наличной реакции добавляется и следовая (длительность стимула равна 4 с); но уже второе предъявление раздражителя вызывает подавление альфа-ритма, заканчивающееся в среднем уже в течение 3 с, т. е. еще до завершения звукового раздражителя. Примерно с 8 – 9‑го применения стимула длительность реакции является минимальной, составляя в среднем около 0,5 с и менее.
В целом кривая угашения ЭЭГ ориентировки весьма напоминает гиперболу (следует иметь в виду, что она, как и некоторые предыдущие, построена в предположении нулевых реакций у тех испытуемых, которые достигли критерия раньше, чем за 20 предъявлений звукового стимула). Однако эта почти правильная функция слагается из индивидуальных кривых, которые демонстрируют весьма широкий диапазон различий от испытуемого к испытуемому и от реакции к реакции. По нашим данным, угашение ЭЭГ ориентировки на звук требует от одного до нескольких десятков применений раздражителя (в среднем около 9), а длительность реакции на первое предъявление стимула составляет от нуля до 26 с (в среднем около 6 с).
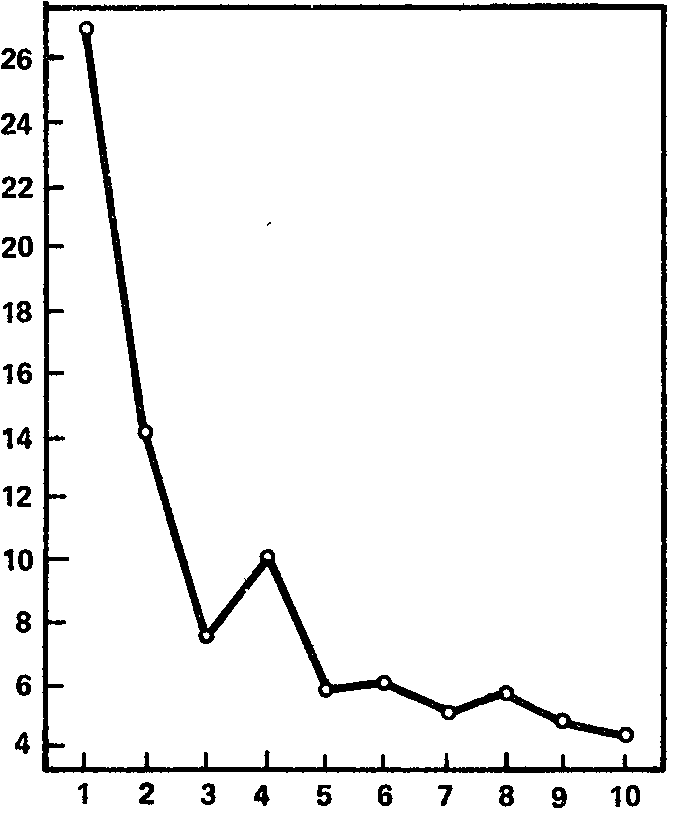
Рис. 16. Динамика ориентировочной электрокорковой реакции на свет (в среднем для всей выборки). Ось абсцисс – порядковый номер светового раздражителя; ось ординат – длительность реакции десинх-ронизации (в с).
На рис. 16 показана динамика ориентировочной реакции биотоков мозга на раздражитель другой модальности, именно на световой стимул значительной яркости (1000 лк при измерении освещенности в непосредственной близости к поверхности излучателя). Измерялось следовое действие раздражителя, т. е. длительность блокады альфа-ритма с момента прекращения светового стимула и до восстановления альфа-ритма (группы альфа-волн длительностью не менее 1 с). Как видно из графика, построенного по данным 20 испытуемых, длительность десинхронизации при первом предъявлении света составляет около 27 с, что примерно в 4 раза превышает длительность реакции на первый звук; подобное же соотношение сохраняется и для каждого из последующих 9 применений светового раздражителя. Однако по своей форме кривая сокращения реакции на свет очень близко напоминает кривую, полученную для звукового раздражителя; это и дает основания думать, что, по крайней мере, следовая десинхронизация на световой раздражитель является по своим механизмам собственно ориентировочной реакцией.
Что касается индивидуальных особенностей реакции ЭЭГ на световой стимул, то они так же разнообразны, как и при действии звукового раздражителя. На первое применение света продолжительность следовой блокады альфа-ритма, по полученным нами данным, составляет от 0,6 до 89,0 с, средняя длительность следовой блокады из 10 предъявлений варьирует у разных испытуемых от 0,8 до 31,1 с, составляя в среднем 8,9 с. Индивидуальные кривые, отражающие динамику сокращения следовой блокады альфа-ритма, показывают, что процесс этот совершается подчас с весьма значительными колебаниями эффекта от стимула к стимулу.
По мнению некоторых авторов (А. Юс, К. Юс, 1954; S. Sharpless, Н. Jasper, 1956; Е.Н. Соколов, 1958 а; О.С. Виноградова, 1961), сокращение активации до длительностей, меньших, чем продолжительность действия раздражителя, связано с переводом активирующих механизмов с мезэнцефалического на таламический уровень: первый обеспечивает длительную, но быстро угашаемую тоническую форму реакции, а второй – более короткую, но и медленнее угашаемую фазическую форму реакции активации.
Признание этой точки зрения необходимо ведет к следующим заключениям: 1) во-первых, нужно согласиться с тем, что активация того или другого уровня, по-видимому, зависит от сенсорной модальности раздражителя, так как при звуковом стимуле переход от тонической активации к фазической совершается (в среднем) уже со второго предъявления (А. Юс, К. Юс, 1958), а при световой стимуляции такого перехода может не наблюдаться даже после многих десятков предъявлений; 2) во-вторых, анализируя индивидуальные данные, следует, очевидно, допустить, что различные индивиды могут характеризоваться различным соотношением двух указанных уровней активации: ведь некоторые из них с первого же предъявления стимула демонстрируют фазическую реакцию, у других же даже при звуковом раздражителе трансформация тонической реакции в фазическую требует нескольких предъявлений.
К сожалению, мы не располагаем материалом, который позволил бы подтвердить или опровергнуть оба заключения; между тем, по крайней мере, второе из них, несомненно, представляет интерес для психофизиологии индивидуальных различий, давая возможность в качестве гипотезы выделить лиц с явно таламическим характером неспецифических активирующих воздействий и лиц, нервная система которых имеет склонность к более генерализованной (топографически и во времени) активации, имеющей преимущественно мезенцефалическое происхождение.
Итак, мы располагаем рядом показателей ЭЭГ ориентировки, полученных в двух сериях экспериментов. Каковы соотношения этих показателей между собой? В табл. 44, помещенной в гл. XI и обсуждаемой там по другому поводу, представлены интеркорреляции данных второй серии (20 испытуемых). Данные этой таблицы свидетельствуют о существовании следующих значимых соотношений: между длительностью первой реакции на звук и длительностью угашения ориентировки на звук (r = 0,493; р < 0,05), а также средней длительностью реакции на свет (в 10 предъявлениях; r = 0,447; р < 0,05); между длительностью угашения ориентировки на звук и длительностью реакции на первое предъявление света (r = 0,665; р < 0,01), а также средней длительностью реакции на световой стимул (r = 0,700; р < 0,001); между длительностью первой реакции на свет и средней длительностью реакции на световой раздражитель (в 10 предъявлениях; r = 0,855; р < 0,001). Иными словами, все соотношения между показателями ЭЭГ ориентировки (кроме одного: между длительностями первых реакций на звук и на свет) оказались статистически значимыми, но заметим, одни – на более высоких, другие – на гораздо более низких уровнях вероятностей нулевой гипотезы. Последнее заставляет предполагать наряду с возможным существованием общего фактора, связанного с протеканием ориентировочных реакций наличие некоторых более дифференцированных влияний на различные параметры ЭЭГ ориентировки, связанных как с модальностью применяемых раздражителей, так и с изучаемой функцией.
Это предположение становится еще более обоснованным, когда мы начинаем сравнивать параметры ориентировочной реакции с некоторыми референтными индикаторами свойств нервной системы. В наших сопоставлениях (В.Д. Небылицын, 1963 б, 1964 в; В.Д. Небылицын и др., 1965) мы имели возможность сравнить показатели ЭЭГ ориентировки, во-первых, с некоторыми индикаторами динамичности, полученными внутри той же электроэнцефалографической методики, а во-вторых, с рядом показателей силы нервной системы по отношению к возбуждению. Заметим сразу, что сколько-нибудь значимой связи между параметрами ориентировки и силой нервной системы нам найти не удалось. Правда, в некоторых случаях длительность угашения ориентировки на звук обнаруживала слабую положительную связь с показателями силы (r = 0,20 – 0,30), но корреляции ни разу не достигали хотя бы 10‑процентного уровня значимости.
Что же касается связи параметров ЭЭГ ориентировки с показателями динамичности, то здесь заслуживают быть отмеченными следующие моменты (табл. 44):
1) длительность реакции десинхронизации на первое предъявление звукового стимула высоко коррелирует со средним условно-рефлекторным эффектом, полученным при «нейтральном» подкреплении (r = 0,709; р < 0,001), в то время как коэффициенты корреляции этого показателя с другими ЭЭГ индикаторами не превышают 0,45. Это – данные второй нашей электроэнцефалографической серии. В первой серии соотношение коэффициентов было примерно таким же, хотя некоторые коэффициенты были несколько выше; корреляция рангов между длительностью первой реакции и средними условнорефлекторным эффектом составила 0,72 (р < 0,001), а для средней длительности условий реакции, выработанной на активирующем подкреплении, – 0,62 (р < 0,01);
2) скорость угашения ориентировки (точнее, число применений стимула до достижения критерия) имеет значимую корреляцию со скоростью угашения условной реакции, выработанной на активирующем подкреплении (r = 0,600; р < 0,01); в то же время корреляция этого показателя со средним условнорефлекторным эффектом не достигает даже 10 %-ного уровня значимости (r = 0,358; р > 0,1). В первой серии соответствующие корреляции были значительно выше – соответственно 0,76 (р < 0,001) и 0,66 (р < 0,01), но между угашением ориентировки и средней длительностью условной реакции, образованной на активирующем подкреплении, корреляция была близка к нулю (0,04);
3) средняя (из 10 предъявлений светового раздражителя) длительность следового подавления альфа-ритма, которую можно рассматривать как индикатор, в определенной степени отражающий длительность угашения ориентировки на свет (чем больше средняя следовая десинхронизация, тем дольше, следовательно, длится процесс привыкания к стимулу), высоко коррелирует с длительностью угашения условной десинхронизации (г = 0,687; р < 0,001); корреляция этого же показателя со средним условным эффектом гораздо ниже и находится на грани значимости (г = 0,449; р < 0,05). В первой серии аналогичный показатель отсутствовал;
4) наконец, длительность блокады альфа-ритма при первом предъявлении света тоже имеет значимую корреляцию с показателем угашения условной реакции (г = 0,622; р < 0,01) и гораздо более низкую – с показателем среднего условного эффекта (r = 0,334; р > 0,1). В первой серии аналогичные корреляции были значимыми (соответственно 0,58, р < 0,01 и 0,69, р < 0,01).
Приведенные соотношения по своему характеру и смыслу близко напоминают те, которые были вскрыты при рассмотрении данных кожно-гальванической методики. Действительно, главный вывод, следующий из анализа таблицы интеркорреляций, заключается в данном контексте в том, что быстрота угашения ориентировочной ЭЭГ реакции на звук и на свет очень хорошо коррелирует с референтным показателем динамичности тормозного процесса – скоростью угашения условной реакции, но довольно слабо (в одном случае – незначимо) коррелирует с референтным показателем динамичности возбуждения – средним условнорефлекторным эффектом. Что же касается длительностей реакции десинхронизации на первые предъявления звукового и светового раздражителей, то их соотношения, как и в случае КГР, выглядят более сложно обусловленными, а кроме того, как видно, зависят от сенсорной модальности стимулов. Если реакция на первое предъявление звука, являющегося нейтральным стимулом для ЭЭГ, высоко коррелирует со средним условным эффектом и слабо – с длительностью угашения условной реакции, то для реакции на первое предъявление света, являющегося специфическим депрессором альфа-ритма, эти соотношения являются, скорее, обратными. Ниже, в гл. XI, при изложении результатов факторного анализа интеркорреляций, представленных в табл. 44, возможные причины этих расхождений будут обсуждены более подробно.
Завершая рассмотрение изложенного в данной главе материала, мы приходим к выводу о том, что параметры любого по физиологической модальности ориентировочного рефлекса организованы в достаточно сложную функциональную структуру, отдельные элементы которой испытывают влияния со стороны различных свойств нервной системы. Так, можно с большой долей уверенности утверждать, что быстрота угашения ориентировочных реакций определяется главным образом динамичностью тормозного процесса как свойством, связанным вообще с быстротой генерации потенциала торможения.
Об этом говорят особенно убедительно результаты, полученные кожно-гальванической и электроэнцефалографической методиками, – высокие корреляции между показателями угашения ориентировки на нейтральный звуковой раздражитель и показателями выработки дифференцировки и угашения условной реакции. Это заключение подтверждается и негативными результатами – слабыми и незначительными корреляциями между угашением ориентировки и образованием положительных условных реакций, зависящим от динамичности возбудительного процесса.
Можно, далее, заключить, что величина реакции на первое предъявление стимула испытывает на себе двойственное влияние как со стороны динамичности возбуждения, так и главным образом со стороны динамичности торможения: первое свойство, видимо, определяет размах, амплитуду, длительность реакции, однако без вмешательства тормозного процесса реакция, возможно, продолжалась бы неограниченно долго. Тормозный процесс выступает в роли ограничителя реакции, и момент его введения в качестве такового определяется как раз уровнем его динамичности. Таким образом, взаимодействие тормозных влияний с возбудительными формирует функцию, обладающую свойством автостабилизации, при этом быстрота развития стабилизирующего эффекта зависит от динамичности тормозного процесса. Конечно, эта схема носит достаточно умозрительный характер, практически же отделить влияние динамичности возбуждения от динамичности торможения в протекании каждой отдельной ориентировочной реакции невозможно.
Далее, в некоторых параметрах ориентировочной реакции можно предполагать влияние силы нервной системы (относительно возбуждения). Это влияние экспериментально показано при изучении индивидуальных различий в направлении сенсорных ориентировочных реакций, и его можно также с определенностью предполагать в величине порога ориентировочной реакции. Нам не удалось выявить отчетливого влияния силы нервной системы в амплитуде ориентировочной реакции, хотя некоторые указания на существование такой зависимости в сенсорных ориентировочных реакциях все же были получены. Скорее всего, влияние силы (чувствительности), если оно имеется, перекрывается влиянием других факторов деятельности нервной системы, более эффективных в отношении данной функции, именно динамичности торможения и динамичности возбуждения. Эти два свойства нервной системы, особенно первое, видимо, являются главными детерминантами основных динамических параметров ориентировочного рефлекса – его величины и скорости его угашения.
Глава 5. Проявления динамичности нервной системы в некоторых электроэнцефалографических показателях
Как мы могли уже убедиться, электроэнцефалография предоставляет в распоряжение исследователя ряд удобных индикаторов основных свойств нервной системы, и в частности динамичности нервных процессов, основанных на изучении феномена активации, т. е. блокады, подавления доминирующего ритма.
Единственным, но, к сожалению, непреодолимым ограничением этого метода, препятствующим его широкому применению для изучения индивидуальных особенностей, является отсутствие достаточно четко выраженного альфа-ритма у части нормальных взрослых индивидов (примерно у 15 – 20 %) и, следовательно, невозможность применения к ним методов, основанных на определении параметров ориентировочной и условнорефлекторной реакций активации.
Это существенное препятствие к широкому использованию ЭЭГ методики может быть преодолено, если будут отысканы такие электроэнцефалографические индикаторы, которые не сводятся к параметрам реакции активации и сами по себе отражают те или иные свойства нервной системы.
В данной главе излагаются некоторые предварительные результаты попыток решить эту задачу.
Мы рассмотрим здесь два вопроса.
Один из них касается соотношения между динамичностью нервных процессов и показателями ЭЭГ покоя, или «фоновыми» показателями ЭЭГ, а другой – соотношения между динамичностью нервных процессов и индикаторами одной из наиболее часто применяемых «функциональных нагрузок», а именно реакции навязывания ритма мелькающего светового раздражителя.
|
|
|
|
|
Дата добавления: 2015-06-26; Просмотров: 429; Нарушение авторских прав?; Мы поможем в написании вашей работы!