КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Г. Электромиографические индикаторы динамичности тормозного процесса 1 страница
|
|
|
|
Специфическим аспектом проблемы динамичности тормозного процесса является вопрос о показателях этого свойства, извлекаемых из электромиографической методики. В отличие от описанных выше фотохимических, электрокорковых и кожно-гальванических реакций мышечная функция является вполне «произвольной» – двигательная активность поперечнополосатой мускулатуры легко поддается достаточно тонкому сознательному регулированию и управлению. При исследовании у нормального человека двигательных реакций, обусловленных предварительной инструкцией, эта произвольность выражается в формировании «с места» любых не слишком сложных систем связей в двигательном анализаторе, а при электромиографической регистрации – в незамедлительном появлении импульсной мышечной активности при осуществлении действий, обусловленных инструкцией. В силу этого обстоятельства и двигательная методика с предварительной инструкцией и ее электромиографическое развитие, видимо, не могут быть применены для изучения динамичности возбудительного процесса. Показатели этого свойства средствами двигательной методики практически неуловимы.
Совсем по-иному, однако, обстоит дело с индикаторами динамичности торможения. Хотя произвольное, совершаемое согласно инструкции прекращение, затормаживание двигательных реакций как таковых опять-таки не представляет для испытуемых каких-либо затруднений, оказывается, что затормозить с такой же легкостью электрическую импульсную активность мышц могут отнюдь не при всех условиях и не все испытуемые. «…Человек может произвольно воздержаться от совершения движения, но он не может произвольно прекратить электрическую активность мышц при наличии даже самой слабой тенденции к совершению движения» (Б.М. Теплов, 1963, с. 16). Эта особенность электромиографической методики и открывает путь к использованию предоставляемых ею показателей в качестве индикаторов динамичности торможения, поскольку индивидуальные различия в легкости и быстроте подавления электрической мышечной активности вне моментов собственно двигательной деятельности, с нашей точки зрения, ближе всего толкуются именно как проявления различного уровня динамичности тормозного процесса.
Проблема индивидуальных различий, вскрываемых электромиографической методикой, была тщательно в целом ряде экспериментальных серий изучена А.Я. Колодной (1959, 1961, 1963 а, б). Основным методическим приемом служило в этих работах предъявление ритмической световой стимуляции (одна вспышка в 1,2 с) с задачей испытуемому нажимать на резиновый баллончик одной или двумя руками в порядке, установленном для данного опыта инструкцией. Различные варианты экспериментальной задачи позволяли извлечь из опыта несколько количественных показателей, таких, например, как длительность подготовительной активности мышц при экстренном увеличении периода между сигналом «Внимание!» и включением ритмического раздражителя или число опытов до выработки так называемых чистых пауз, т. е. интервалов между двумя нажатиями, свободных от электрических мышечных импульсов, и некоторые другие.
Рисунки 12 и 13 иллюстрируют индивидуальные различия в элект-ромиограммах локтевых сгибателей кисти в условиях заданного инструкцией чередования одного положительного и нескольких (двух и пяти) тормозных раздражителей. Из рисунков видно, что, хотя все испытуемые исправно выполняют инструкцию, нажимая только в ответ на положительные сигналы, в динамике мышечной электрической активности наблюдаются разительные индивидуальные вариации. Если у некоторых испытуемых электрические ирияв-ления мышечной активности хорошо соответствуют по времени наличию положительного раздражителя, то у других испытуемых мышечной активностью заполнены и паузы между положительными сигналами. Столь же существенные различия наблюдаются и при регистрации подготовительной активности мышц, а также при записи мышечных импульсов по окончании серии положительных раздражителей – в «последействии» ритмической последовательности сигналов.

Рис. 12. Электромиографическая активность сгибателей кисти у двух испытуемых с малой динамичностью тормозного процесса: А – при чередовании одного положительного сигнала с двумя тормозными; Б – при чередовании одного положительного и пяти тормозных сигналов. 1 – отмегка положительной реакции, 2 – ЭМГ, 3 – отметка световых сигналов, 4 – отметка времени (1 с) (А.Я. Колодная, 1961).
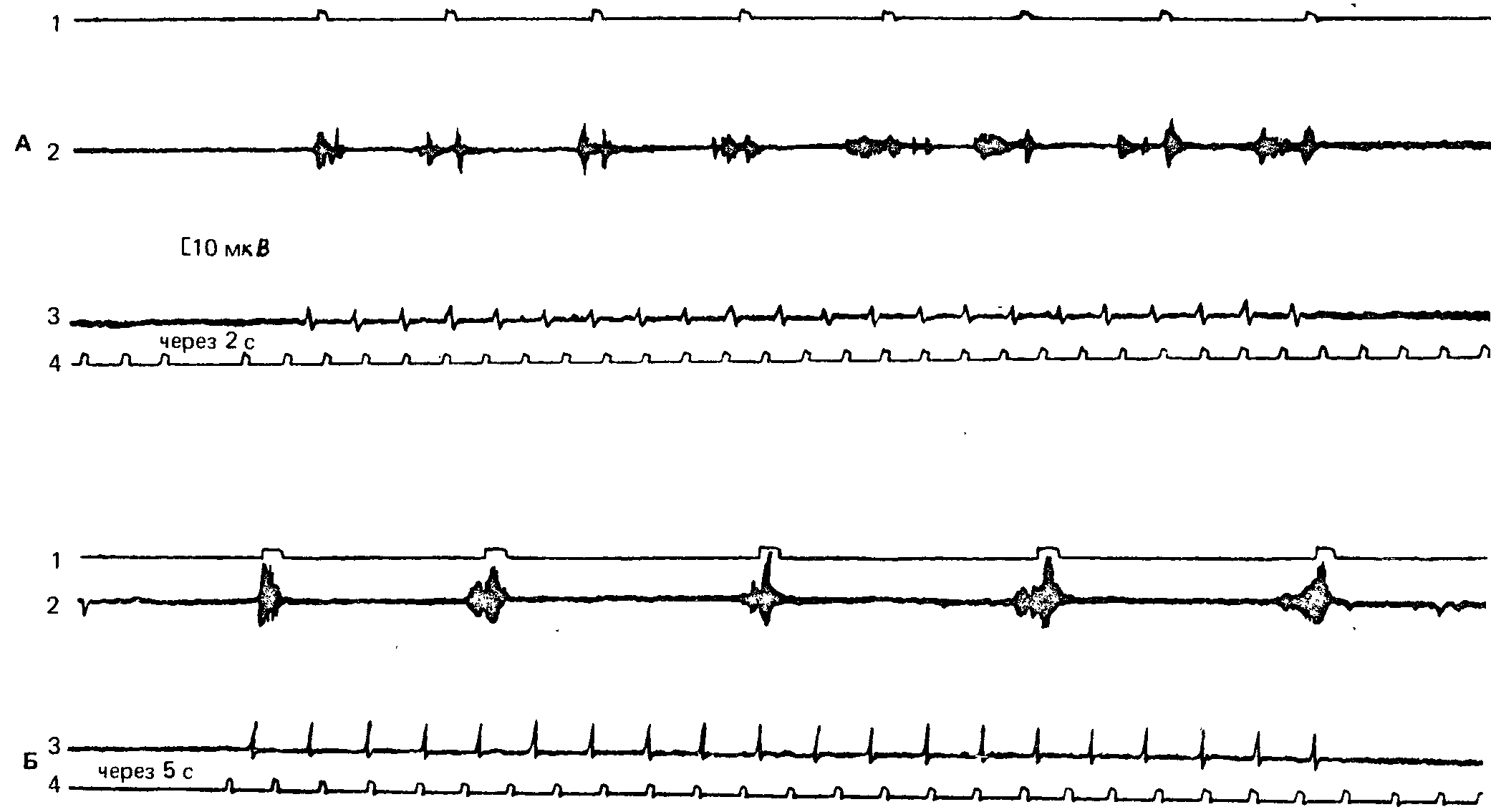
Рис. 13. Электромиографическая активность сгибателей кисти у двух испытуемых с высокой динамичностью тормозного процесса. Обозначения те же, что на рис. 12 (А.Я. Колодная, 1961).
По данным А.Я. Колодной (1963 а), число предъявлений ритмической серии стимулов до выработки чистых пауз варьирует у разных испытуемых от 3 до 390, процентное отношение чистых пауз к общему числу интервалов – от 98 до нуля, время, занятое мышечной активностью в интервале экстренного отставления стимуляции на 20 с, – от 0,2 до 20 с и, наконец, длительность последействия по прекращении стимуляции – от нуля до 17 с, при переходе от одной руки к другой – от нуля до 31 с.
Каждый из указанных показателей может быть с достаточными основаниями интерпретирован как индикатор динамичности тормозного процесса. Это не вызывает сомнений по отношению к такому показателю, как скорость выработки чистых пауз. Совершенно очевидно, что успех этой процедуры зависит в полной мере от легкости и быстроты генерации нервной системой процесса торможения в ходе многократного предъявления стимуляции, обладающей тормозным значением. Эта процедура в некотором смысле аналогична выработке дифференцировки. Но и такой показатель, как импульсная активность в период неожиданного отставления стимуляции, видимо, тоже определяется динамичностью торможения, поскольку для подавления этой активности требуется экстренное развитие тормозного процесса. Наконец, от динамичности торможения, как можно думать, в значительной степени зависит и прекращение мышечных импульсов в последействии, вызванном ритмической стимуляцией: здесь тормозный процесс играет роль активного ограничителя следовых явлений, которые без его вмешательства, возможно, продолжались бы неопределенно долгое время.
Представление о том, что во всех этих индикаторах наблюдается действие единого физиологического фактора, которым, вероятно, является, динамичность тормозного процесса, подтверждается их взаимными сопоставлениями как в форме простого сравнения результатов отдельных испытуемых, так и с помощью корреляционного анализа.
Таблица 1
Интеркорреляции рангов между электромиографическими показателями динамичности тормозного процесса (М.Н. Борисова и др., 1963)
| Показатели | ||||||
| 1. Время, занятое мышечной активностью в интервале 4 с отставления стимуляции | ||||||
| 2. То же в интервале 20 с отставления | ||||||
| 3. Длительность мышечного последействия по прекращении стимуляции | ||||||
| 4. Сохранение мышечной активности | ||||||
| в неработающей руке при переключении на другую руку | ||||||
| 5. Отношение числа чистых пауз к общему числу пауз между стимулами | ||||||
| 6. Число опытов до выработки чистых пауз |
Примечания. 1. В этой и во всех последующих корреляционных таблицах нули и запятые у числовых значений коэффициентов корреляции опущены, даны только десятичные знаки. 2. Все коэффициенты достоверны на 0,1 %-ном уровне значимости.
В табл. 1 приведены интеркорреляции рангов шести электромиографических показателей, использованных в числе прочих для сопоставления в коллективной работе М.Н. Борисовой с соавторами (1963). Высокие числовые значения каждого из коэффициента (р < 0,001) свидетельствуют о наличии в матрице интеркорреляций общего фактора, который действительно был выделен авторами указанной работы. Правда, он был интерпретирован как баланс нервных процессов, т. е. как соотношение двух процессов – возбуждения и торможения, однако толкование его с точки зрения функции одного процесса, именно торможения, кажется более экономным и рациональным. Оно позволяет, в частности, избежать вынужденного ограничения, вводимого интерпретацией этого фактора как баланса нервных процессов в характеристику испытуемых, различающихся по уровню мышечной импульсной активности. Дело в том, что с точки зрения баланса эти испытуемые – если взять крайние группы – характеризуются либо как возбудимые, т. е. с преобладанием возбуждения, либо как уравновешенные. В последнюю категорию попадают лица с минимальной мышечной активностью (А.Я. Колодная, 1961, 1963 б). Как видим, при таком подходе не могут быть выделены в особую группу индивиды с преобладанием тормозного процесса – для них просто не предусматривается места. Если же мышечную активность интерпретировать как функцию динамичности торможения, то тогда указанная трудность отпадает: значительная мышечная активность будет признаком низкой, а малая активность – высокой динамичности тормозного процесса. Таким образом, весь континуум этого свойства оказывается полностью занят распределением данной физиологической функции, и ни один из его полюсов не остается не заполненным тем или иным функциональным содержанием, как это имеет место при истолковании электромиографических показателей с позиции баланса нервных процессов.
Возникает вопрос: каковы внутренние мозговые механизмы генерации тормозного потенциала при выработке внутреннего торможения? И более частная проблема: одни ли и те же тормозные механизмы принимают участие в формировании рассмотренных выше разновидностей внутреннего торможения – дифференцировочного и угасательного, или эти механизмы различны и, таким образом, могут не совпадать у одного и того же испытуемого?
Ответ на первый вопрос в принципе может быть двояким. Выше уже указывалось, что современная нейрофизиология допускает возникновение тормозного состояния в данной структуре или системе двумя путями: либо путем внутренней трансформации состояния данной структуры, бывшей непосредственно перед этим в состоянии возбуждения, либо путем активации специальных тормозных экстраструктур, посылающих при определенных условиях тормозные команды. Имеются свидетельства о существовании ряда таких тормозящих образований, действующих в качестве ингибиторов относительно моторики, примитивных эмоций, вегетативных реакций, т. е. в качестве генераторов, главным образом безусловных тормозных приказов. Можно думать, однако, что в выработке условного торможения ведущую роль играют если не все, то, по крайней мере, некоторые отделы верховного регулятора биологических функций – коры больших полушарий. Эта точка зрения была подробно обоснована нами в статье, специально посвященной проблеме нейрофизиологических механизмов баланса нервной системы по динамичности возбудительного и тормозного процессов (В.Д. Небылицын, 1964 б).
Что касается вопроса о совпадении или несовпадении механизмов дифференцировочного и угасательного торможения то, исходя из только что сформулированного представления, следует, очевидно, предположить, что эти механизмы должны быть, – по крайней мере, в общем – теми же самыми как в случае дифференцировки, так и в случае угашения. К сожалению, мы располагаем слишком незначительным экспериментальным материалом для прямого ответа на этот вопрос. Все же те сведения, которые имеются, говорят, скорее, в пользу этого предположения.
Так, напомним высокое соответствие между данными, полученными Н.И. Майзель (1956) по фотохимической методике. В наших опытах с использованием ЭЭГ показателей мы имели возможность сравнить выработку электрокорковой дифференцировки с угашением положительной условной реакции. Коэффициент корреляции рангов, если сравнивать испытуемых по скорости достижения критерия, был не очень высоким (0,42; р < 0,05); если взять другой показатель, именно среднюю длительность десинхронизации в первых 5 пробах тормозного стимула, то корреляция рангов возрастает до 0,57 (р < 0,02). Эти не слишком высокие показатели связи хотя и указывают на общность механизмов внутреннего торможения (по крайней мере, при одном и том же эффекторе), но наводят в то же время на мысль о том, что на фоне совпадения этих функций все же, по-видимому, существует и известная их независимость, проявляющаяся иногда в расхождении скоростей формирования этих двух видов внутреннего торможения. В нашем электроэнцефалографическом материале такое расхождение мы наблюдали у нескольких испытуемых: у одного из них быстро образовывалась дифференцировка (9 сочетаний до выработки), но не удалось добиться угашения условной реакции, а у трех других имело место обратное соотношение (угашение после 3, 3 и 7 предъявлений и выработка дифференцировки соответственно после 35, 52 и 69 сочетаний, перемежающихся через 2 – 4 сочетания предъявлением дифференцировочного стимула) (В.Д. Небылицын, 1963 б).
Обсуждая возможный физиологический смысл подобных расхождений, можно думать, что, хотя кора головного мозга и осуществляет ведущую тормозную функцию, тем не менее другие (вероятно, подчиненные ей) тормозящие образования могут в некоторых ситуациях выработки внутреннего торможения играть более значительную роль. Так, если принять гипотезу А.И. Ройтбака (1962), связывающую угашение условных рефлексов с активацией неспецифической системы таламуса, то можно предположить, что в процессе угашения, в отличие от выработки дифференцировки, неспецифический («ретикулярный») таламус может в каких-то случаях обеспечивать сравнительно более быстрое развитие торможения. Эти случаи, возможно, и являются причиной понижения коэффициента корреляции между скоростью образования дифференцировки и быстротой угашения условных рефлексов.
Глава 4. Ориентировочные реакции и зависимость их динамики от основных свойств нервной системы
Реакция животного на новизну была впервые исследована и получила название ориентировочного рефлекса в школе И.П. Павлова. Было показано, что возникновение ориентировочного рефлекса не связано с сенсорной модальностью раздражителя, что он может быть подвергнут угашению, причем механизмом последнего является генерация внутреннего торможения, что при всем том он является врожденным, т. е. безусловным, и сохраняется у животных, лишенных коры больших полушарий, приобретая в этом случае особую стойкость и неугасимость (Н.А. Попов, 1921, 1938; С.Н. Чечулин, 1923; И.С. Розенталь, 1929; Г.П. Зеленый, 1930).
Первоначально ориентировочным рефлексом называлась только двигательная реакция животного в сторону нового или необычного раздражителя (поворот головы, движение ушами и глазами и т. д.). Впоследствии получила распространение более широкая точка зрения, согласно которой ориентировочный рефлекс представляет собой целую систему реакций, интегрированных в сложном соматове-гетативном комплексе (Е.Н. Соколов, 1958 а, б; О.С. Виноградова, 1959, 1961). Таким образом, ориентировочная реакция может быть изучена как по моторным, так и по вегетативным и электрографическим индикаторам, которые, впрочем, не всегда согласуются друг с другом (например, скорость угашения различных компонентов ориентировочной реакции может быть различна у одного и того же испытуемого).
Ориентировочная реакция может быть охарактеризована целым рядом параметров, каждый из которых имеет особое функциональное значение, по-видимому, не всегда совпадающее со значением других. Относительно каждого из них можно предполагать ту или иную степень связи с определенными особенностями нервной системы. Каковы эти параметры?
Один из них – порог ориентировочного рефлекса. Поскольку последний всегда есть результат сенсорной стимуляции, постольку возникает вопрос о том минимальном значении стимула, которое вызывает ответ в виде ориентировочной реакции. Многими авторами было установлено, что порог ориентировочного рефлекса (в основном по кожно-гальваническому и электроэнцефалографическому показателям) фактически совпадает с порогом ощущения, определяемым словесной реакцией, во всяком случае, до того, как ориентировочная реакция начинает угасать при повторном предъявлении стимула (Г.В. Гершуни, 1955; A.J. Derbyshire, J. С. Farley, 1959). Но порог ощущения (подробнее см. об этом ниже) обнаруживает связь с силой нервной системы (Б.М. Теплов, 1955; В.Д. Небылицын, 1959 а; В.И. Рождественская и др., 1960). Следовательно, порог возникновения ориентировочной реакции должен коррелировать с показателями силы нервной системы (относительно возбуждения).
К сожалению, до сих пор прямого сопоставления соответствующих индикаторов в эксперименте не было приведено, хотя, вероятно, использование этого приема принесло бы пользу при изучении соотношения между чувствительностью и силой нервной системы у животных.
В типологическом контексте может исследоваться и другой параметр ориентировочной реакции – ее величина. Определение этого параметра представляет некоторые трудности, так как величина ориентировочной реакции закономерно падает по мере повторения предъявлений. Поэтому для учета величины ориентировочного рефлекса нужно пользоваться одним из следующих приближенно отвечающих задаче показателей: 1) величиной реакции на первое предъявление нового стимула, 2) средней величиной реакции на некоторое заранее фиксированное число предъявлений стимула, наконец, 3) характеристикой крутизны кривой, отображающей на графике динамику угасания ориентировочной реакции (градиент функции). Самый простой из этих показателей – первый, и, как мы увидим впоследствии, он достаточно хорошо себя проявляет.
Наконец, третий основной параметр ориентировочной реакции – это скорость угашения ее при продолжающемся повторении стимула. Угашение может производиться до определенного, заранее намеченного критерия, например до отсутствия реакции в серии из трех или более предъявлений подряд (острое угашение) или до отсутствия реакций в нескольких следующих друг за другом опытах (хроническое угашение). Эта процедура сильно напоминает угашение условного рефлекса. И.П. Павлов предполагал, что она тоже сопровождается развитием внутреннего торможения (1951–1952, т. IV, с. 269) и, возможно, по физиологическому смыслу означает то же самое, что и угашение условной реакции. Поскольку, однако, ориентировочный рефлекс есть безусловная реакция, многие зарубежные авторы предпочитают вместо термина «угашение» пользоваться терминами «привыкание» и «адаптация».
Как уже было сказано, каждый из перечисленных основных параметров ориентировочной реакции, вероятно, имеет типологическое значение, т. е. находится в зависимости от каких-либо свойств нервной системы. К сожалению, в павловской школе – как при жизни И.П. Павлова, так и после его смерти – не были проведены сколько-нибудь систематические исследования индивидуальных особенностей ориентировочных реакций, равно как и возможной связи этих особенностей со свойствами нервной системы, хотя данные, полученные попутно некоторыми из упомянутых выше авторов, несомненно, давали основания думать, что в ряде особенностей динамики ориентировочного рефлекса отражаются и свойства нервной системы животного. Имеющиеся прямые данные по сопоставлению свойств ориентировочной реакции со свойствами нервной системы могут быть систематизированы следующим образом.
В 1933 г. Н.В. Виноградовым была описана собака слабого типа, которая, по наблюдениям автора, характеризовалась неугасимым ориентировочным рефлексом. С тех пор в литературе (М.С. Колесников, 1953) существует мнение о том, что животным слабого типа нервной системы свойственна неугасающая ориентировочная реакция на любые раздражители внешней среды. Таким образом, согласно этой точке зрения, скорость угашения ориентировки есть функция силы нервной системы.
Другая точка зрения (Л.Н. Стельмах, 1956) связывает быстроту угашения ориентировочной реакции не с силой нервной системы, а с подвижностью нервных процессов (определяемой скоростью переделки). Л.Н. Стельмах указывает, что, с одной стороны, неугасимая ориентировочная реакция может встречаться и у собак сильного типа, а с другой стороны, угашение ориентировки может быть легко достигнуто и у собак со слабой нервной системой. В то же время обнаруживается некоторая зависимость быстроты угашения от свойства подвижности (правда, с существенными исключениями). К сожалению, автор не приводит количественных значений связи между угашением ориентировки и переделкой. Существенным недостатком работы является также то, что исследование ориентировочной реакции было проведено уже после того, как у собак был определен тип нервной системы, т. е. после многомесячной работы с разнообразными внешними раздражителями.
Э.А. Варуха (1953), сопоставляя динамику ориентировочных реакций у собак с результатами определения свойств нервной системы по малому стандарту, нашел, что такой показатель, как изменение величины ориентировочного рефлекса при усилении раздражителя, может быть взят для оценки силы нервной системы (относительно возбуждения), а быстрота угашения ориентировки не связана с силой нервной системы относительно торможения.
Работы, выполненные Л.Г. Ворониным, Е.Н. Соколовым и их сотрудниками (Л.Г. Воронин, Г.И. Ширкова, 1949; Л.Г. Воронин, Е.Н. Соколов, 1955; Е.Н. Соколов и др., 1955; Л.Г. Воронин и др., 1959; У. Бао-Хуа, 1958, 1959), обратили внимание на другой аспект типологической обусловленности ориентировочных реакций, а именно на связь их с уравновешенностью нервных процессов. При этом, как уже указывалось в гл. II, хотя авторы говорят об уравновешенности по силе, анализ применяемых ими испытаний позволяет заключить, что речь идет, скорее, о том, что мы обозначаем как уравновешенность нервных процессов по динамичности. Так, в работе У. Бао-Хуа (1959) референтным показателем уравновешенности служило количество ошибочных действий при выработке элементарного двигательного стереотипа по предварительной инструкции, точнее, соотношение ошибок при предъявлении положительных и отрицательных компонентов стереотипа.
Ни это, ни другие испытания, предусматриваемые методикой Н.А. Рокотовой (1954), примененной в данном случае У. Бао-Хуа, вообще не могут дать показателей силы (выносливости) нервной системы относительно возбуждения, равно как и относительно торможения, но некоторые из них можно трактовать как отражающие уровень динамичности нервных процессов. В большинстве указанных работ речь идет о скорости угашения кожно-гальванических реакций, и предположения сводятся к тому, что быстрое угашение ориентировки по кожно-гальваническому показателю указывает на преобладание тормозного процесса, а замедленное угашение КГР свидетельствует о преобладании возбудительного процесса. То же предположение содержится в работе А. Манди-Касла и Б. Маккивера (А.С. Mundy-Castle, В. Z. МсКiеvеr, 1953), выполненной также с использованием кожно-гальванического показателя.
Итак, разные авторы связывают те или иные показатели ориентировочного рефлекса с различными свойствами нервной системы, причем, как видно, основной интерес вызывает быстрота угашения реакции. Что можно сказать по этому поводу?
Роль силы нервной системы в некоторых характеристиках ориентировочной реакции вряд ли может быть подвергнута сомнению. Мы уже говорили об этом при обсуждении вопроса о пороге возникновения ориентировки. Но величина ориентировочной реакции, по всей видимости, также не может в какой-то степени не зависеть от силы нервной системы относительно возбуждения. Поскольку сильная нервная система обладает менее высокой чувствительностью, зависимость между силой и величиной ориентировки должна быть обратной: индивиды со слабой нервной системой должны обладать более выраженной ориентировочной реакцией, особенно при использовании раздражителей слабой и средней интенсивности, которые в случае систем разной чувствительности обеспечат наибольшие различия в физиологическом эффекте. Возможно, это и является одной из причин более высокой ориентировочной активности, «неугасимого» ориентировочного рефлекса у некоторых особей слабого типа нервной системы – но, вероятно, только одной из причин, причем не самой существенной.
Что касается связи ориентировочных реакций с подвижностью нервных процессов, то имеющиеся материалы (Л.Н. Стельмах, 1956) недостаточны, чтобы сделать по этому вопросу сколько-нибудь определенные выводы. Это, конечно, не означает, что предположение о такой связи должно быть с места отвергнуто. Это означает только, что оно должно быть проверено в экспериментальном сопоставлении соответствующих индикаторов.
Наиболее обоснованными кажутся взгляды, связывающие некоторые параметры ориентировочной реакции с уравновешенностью нервных процессов (мы бы сказали, с уравновешенностью по динамичности). При этом, возможно, следует иметь в виду, что динамичность возбудительного и динамичность тормозного процессов, отражая функционально различные свойства нервного субстрата, могут оказывать различное влияние на разные стороны ориентировочного рефлекса.
Что касается скорости угашения ориентировки, то она, как можно полагать, является прямой функцией динамичности тормозного процесса. Как уже отмечалось, еще И.П. Павлов и его сотрудники указывали, что эффект угашения ориентировочного рефлекса полностью подобен эффекту угашения условного рефлекса: сходство наблюдается как в деталях самих процессов, так и в их результатах – оба они приводят к возникновению дремотного и сонного состояния, обязанного своим происхождением иррадиации вырабатываемого внутреннего торможения.
Анализ электрографических проявлений ориентировочного рефлекса позволил Е.Н. Соколову (1963) и О.С. Виноградовой (1961) выдвинуть предположение о том, что само угашение ориентировочной реакции есть не что иное, как постепенно вырабатываемый условнорефлекторный процесс, в котором условным раздражителем является начало применяемого стимула, становящееся сигналом его определенной длительности и отстутствия его в фоне.
Таким образом, угашение ориентировочного рефлекса ведет к формированию тормозной функциональной структуры так же, как и угашение условной реакции, ведущее, как предполагается, к повышению избирательной активности тормозящих синаптических аппаратов (Е.Н. Соколов, Н.П. Парамонова, 1961; П.В. Симонов, 1962). Так же как и в случае условной реакции, эта тормозная функциональная структура, видимо, складывается прежде всего в коре больших полушарий: удаление коры, согласно данным, полученным еще в школе И.П. Павлова (Г.П. Зеленый, 1930; Н.А. Попов, 1938), и данным новейших работ (М. Jouvet, 1961), приводит к устранению механизма угашения ориентировочной реакции, в результате чего, как указывает Е.Н. Соколов (1963), ориентировочный рефлекс превращается в собственно безусловный рефлекс, лишенный условнорефлекторных компонентов и не поддающийся поэтому угашению.
Основываясь на этих данных и соображениях, мы и предполагаем, что угашение ориентировочной реакции, так же как угашение условной реакции, есть функция главным образом того свойства нервной системы, которое мы обозначаем как динамичность тормозного процесса: высокий уровень динамичности торможения ведет к быстрому угашению ориентировки, при низком уровне этого свойства угашение ориентировки может оказаться весьма длительным процессом. Заметим снова, что последнее явление может, вероятно, быть следствием не только низкой динамичности тормозного процесса, но и высокой абсолютной чувствительности анализатора, воспринимающего сенсорный стимул, который, попадая на данную систему, получает большую физиологическую эффективность; высокая же чувствительность присуща слабой нервной системе.
Некоторые параметры ориентировочной реакции, возможно, зависят и от динамичности возбудительного процесса. В частности, влияние последнего можно предполагать в величине ориентировочной реакции при первых предъявлениях стимула. Действительно, если последующие его предъявления приводят к развитию условного торможения, ограничивающего возникающее возбуждение, то при первом нанесении раздражителя это ограничение еще не выработано или, во всяком случае, недостаточно. Поэтому возбуждение, возникающее при первом предъявлении сигнала, когда механизмы условного торможения еще не вступили в действие, будет, возможно, характеризоваться большей амплитудой, интенсивностью и длительностью. Отсюда у индивидов с высокой динамичностью возбудительного процесса можно ожидать более выраженных (по величине) ориентировочных реакций на первое включение стимула по сравнению с индивидами с низкой динамичностью процесса возбуждения.
По некоторым из высказанных предположений в лаборатории психофизиологии были получены определенные экспериментальные данные. Поскольку эти данные всякий раз имеют свою специфику соответственно использованной методике, рассмотрим их в нескольких разделах, посвящая каждый какой-нибудь одной из примененных методик.
Сенсорные ориентировочные реакции. Специфической чертой сенсорных ориентировочных реакций, т. е. протекающих по правилам ориентировочного рефлекса изменений порогов ощущения (в нашем случае абсолютных порогов), является то, что они кроме указанных выше параметров – порога, величины и скорости угашения – обладают еще и параметром направления: ориентировочная реакция может выражаться как в снижении, так и в повышении абсолютной чувствительности, варьируя в этом своем качестве от испытуемого к испытуемому.
|
|
|
|
|
Дата добавления: 2015-06-26; Просмотров: 430; Нарушение авторских прав?; Мы поможем в написании вашей работы!