КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Блокада в фазу 3 и фазу 4
|
|
|
|
Если деполяризующий импульс приходит в ножку или ветвь пучка преждевременно (т. е. раньше полного исчезновения ее рефрактерности), он может проводиться медленно или полностью блокироваться. Термин «блокада в фазу 3», используемый для обозначения этого явления, просто отражает приход импульса во время фазы 3 потенциала действия. Импульсы, прибывающие по окончании фазы 3, проводятся нормально. Однако если проводящая структура обладает повышенной спонтанной деполяризацией в фазу 4 (обычно вследствие заболевания), она не может удерживать высокий потенциал покоя, позволяя ему смещаться все ближе и ближе к нулевому уровню, в результате чего проводимость этой структуры постепенно снижается. Чем длительнее период диастолы, тем более степень снижения потенциала покоя и выше вероятность нарушения проведения. Блокаду, развивающуюся по такому механизму в конце длительной диастолы, обычно называют «блокадой в фазе 4» [1]. По опыту автора, блок в фазе 4 характерен для внутрижелудочковой проводящей системы и очень редко встречается в АВ-узле. Его возникновение в 13 исследованных случаях обязательно сопровождалось другими признаками поражения системы ножек и ветвей пучка Гиса и не наблюдалось у здоровых лиц. В связи с этим наличие деполяризации в фазу 4 в системе разветвлений пучка Гиса может использоваться как четкое свидетельство патологического процесса в этих структурах. Существование блоков в фазу 3 или фазу 4 в различных частях системы разветвлений пучка Гиса может обусловить появление некоторых из наиболее сложных и необычных форм нарушения проведения. Возможность возникновения блока в фазу 4 при постэкстрасистолических вариантах блокады обсуждалась выше (см. рис. 3.14 и 3.15).
ГЛАВА 4
Аберрантность: электрофизиологические механизмы и электрокардиографические проявления
Д..Y. Сингер и X. К. Коэн (D. H. Singer и Н. С. Cohen)
Форма комплекса QRS зависит от последовательности активации желудочков, причем изменения формы отражают отклонения от нормы при распространении возбуждения. Это может происходить по одной из двух основных причин: 1) смещение места возникновения импульса к желудочковому эктопическому фокусу или к АВ-узловым двойным путям определенного типа; 2) изменение желудочкового распространения наджелудочковых импульсов. Последнее в свою очередь может быть результатом врожденного или приобретенного нарушения проводящей системы. С другой стороны, данное явление может быть функциональным, обусловленным проведением импульса во время рефрактерного периода.
Термины «аберрантность», «аберрация» и «аберрантное желудочковое проведение» с момента их внедрения сэром Томасом Льюисом в- начале нашего столетия [1, 2] применяются в основном при описании функциональных нарушений внутрижелудочкового проведения, особенно тех, которые наблюдаются в сочетании с изменениями длительности сердечного цикла [3—7]. Однако они в равной мере используются и для определения других типов нарушений внутрижелудочкового проведения. В этой главе и в главе 5 термин «аберрантность» будет рассматриваться в широком контексте. Будет также сделан обзор электрофизиологических механизмов, клинических аспектов и роли различных типов нарушений проведения.
Специализированная проводящая система сердца
При рассмотрении проводящей системы ткани сердца удобнее разделить на два основных типа (рис. 4.1, А): 1) рабочий миокард предсердий и желудочков, ответственный за сократимость сердца и появление зубцов Р и комплексов QRS на стандартной ЭКГ; 2) сеть специализированных тканей [8—10] (см. главу 2, том 1), включающую синоатриальный (СА) и атриовентрикулярный (АВ) узлы и разветвления в системе Гис—Пуркинье [10—13]. Описаны также двойные пути в АВ-узле и специализированные предсердные межузловые тракты [9, 14, 15].
Специализированные ткани несут ответственность за нормальное генерирование импульсов и за их быстрое и упорядоченное проведение от места возникновения в другие отделы сердца. Изменения в процессе образования импульсов и (или) в их проведении предшествуют изменениям частоты сердечных сокращений в ответ на меняющиеся физиологические условия, а также многим нарушениям ритма и проводимости. Электрофизиологические характеристики специализированных тканей являются основными детерминантами аберрантности. Активность специализированных тканей не проявляется на стандартной поверхностной ЭКГ из-за их небольшой (относительно рабочего миокарда) массы, что затрудняет электрокардиографическую оценку предшествующих механизмов.
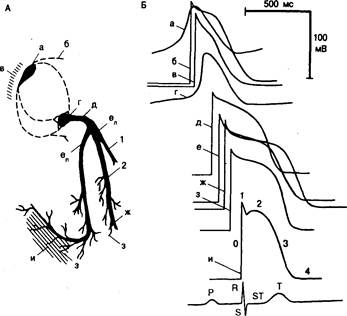
Рис. 4.1. Схематическое изображение специализированной проводящей системы сердца (А) и трансмембранных потенциалов в обычных и специализированных тканях сердца собаки (Б).
Отмечаются различия характеристик потенциалов действия на разных участках сердца. Взаиморасположение зарегистрированных потенциалов отражает последовательность активации. Внизу показана также стандартная поверхностная ЭКГ. Буквами а—и обозначены следующие структуры: а — синусовый узел; б — пучок Брахмана и другие специализированные предсердные межузловые тракты (отмечены пунктирными линиями, соединяющими синусовый и атриовентрикулярный узлы); в — рабочий миокард предсердий; г — атриовентрикулярный узел; д — пучок Гиса; е — ножки пучка; (eп) — правая ножка; (ел) — левая ножка; 1 и 2 — передняя и задняя ветви левой ножки пучка; ж — волокна Пуркинье; з — терминальные волокна Пуркинье в месте их контакта с рабочим миокардом; и — рабочий миокард желудочков. Фазы потенциала действия рабочего миокарда желудочков обозначены арабскими цифрами от 0 до 4, а зубцы на поверхностной ЭКГ — латинскими буквами Р, R, S и Т. Обсуждение в тексте.
Электрофизиологические аспекты
Как отмечалось ранее, аномалии формы комплекса QRS могут возникать по очень многим причинам. В этой главе основное внимание направлено на ряд нарушений, впервые описанных сэром Томасом Льюисом [1, 2], для которых был предложен термин «аберрантность», а именно: аномалии формы комплекса QRS при наджелудочковом возбуждении, наблюдаемые в сочетании с зависимыми от длительности цикла изменениями проводимости и рефрактерности. Аберрантность — это одно из проявлений большой группы зависимых от длительности сердечного цикла нарушений проведения, которая включает в себя такие разновидности, как скрытое проведение [16—18], сверхнормальное проведение [19,20] и связанный с частотой ритма АВ-блок [3—5]. Однонаправленный блок и циркуляция возбуждения [3—7] в некотором смысле также являются проявлениями этого феномена.
Мы рассмотрим аберрантность с точки зрения зависимости изменений комплекса QRS от длительности сердечного цикла, как это предложено Singer и Ten Eick [21]. При ее обсуждении мы будем основываться на корреляции электрокардиографических данных и результатах микроэлектродных исследований трансмембранного потенциала в сердечных тканях животных, а также у больных, подвергшихся операции на открытом сердце.
Электрическая активность сердечных клеток
Понимание механизмов аберрантности зависит от понимания электрофизиологических основ формирования и проведения возбуждения в сердце, а также от знания тех изменений, которые предшествуют развитию нарушений проводимости. Краткий обзор современных концепций будет дан ниже. Для более обстоятельного изучения этого вопроса читателю следует обратиться к главе 3 первого тома данной книги, а также к многочисленным учебникам по физиологии [22—24], обзорам и монографиям по электрофизиологии сердца [25—36].
Нормальный трансмембранный потенциал
Введение стеклянного микроэлектрода [37] в возбудимую клетку позволяет зарегистрировать разность потенциалов между внутренней средой клетки и индифферентным электродом, расположенным снаружи от нее. На рис. 4.1, Б показан идеализированный трансмембранный потенциал, зарегистрированный в разных отделах сердца, вместе с одновременно полученной поверхностной ЭКГ. Фазы потенциала обозначены арабскими цифрами от 0 до 4. Во время электрической диастолы (фаза 4) внутренняя среда клетки отрицательна по отношению к внеклеточной жидкости. В большинстве нормальных сердечных волокон, включая рабочие миокардиальные клетки предсердий и желудочков, разность потенциалов в фазу 4 остается постоянной до момента возникновения возбуждения («потенциал покоя») и в норме составляет в среднем от — 85 до — 95 мВ. При возбуждении в клетке происходит быстрая деполяризация (фаза 0) с кратковременным изменением полярности, после чего начинается процесс постепенной реполяризации (фазы 1, 2 и 3), во время которого мембранный потенциал возвращается к уровню покоя. Сопоставление со стандартной ЭКГ показывает, что фазы 0 и 1 желудочковых потенциалов действия совпадают с зубцами R и S, а фазы 2 и 3 — с сегментом ST и волной Г соответственно (см. рис. 4.1, Б). Трансмембранные потенциалы, зарегистрированные в рабочих и специализированных волокнах различных частей сердца,- могут отличаться друг от друга в некоторых отношениях [25], включая уровень диастолического потенциала, амплитуду потенциала действия, максимальную скорость деполяризации в фазу 0 (Vmax), продолжительность периода реполяризации и длительность потенциала действия (см. рис. 4.1, Б). Отмечаются также различия в возбудимости, проводимости и пейсмекерной способности. Локальные различия электрофизиологических свойств в системе Гис—Пуркинье [25, 31—33] могут иметь важное значение для аберрантности, поскольку они обусловливают локальную вариабельность ответов на различные физиологические и фармакологические факторы [25, 38—47], включающие температуру, рН, частоту, pCO2, гипоксию, ишемию, неорганические катионы и ряд антиаритмических препаратов.
Мембранные механизмы
Электрическая активность сердца является следствием движения ионов через клеточную мембрану [22—36, 48]. Эффект физиологических воздействий и химических веществ, влияющих на образование и проведение импульсов в сердце, обусловлен в основном их способностью изменять ионные токи [24, 30, 37, 47]. На рис. 4.2 показаны основные ионные токи, способствующие развитию потенциала действия в волокнах Пуркинье. Далее мы упрощенно опишем некоторые специфические явления, имеющие отношение к аберрантности.
Мембранный потенциал покоя. Разность потенциала относительно мембраны сердечной клетки обусловлена различием в ионном составе внутриклеточной среды и внеклеточной жидкости [32, 33]. Во внутриклеточной жидкости важнейшим катионом является К+, а доминирующими анионами — фосфаты и радикалы органических кислот. Последние представляют собой в основном поливалентные ионы, часто связанные с белками, для которых клеточная мембрана непроницаема. Во внеклеточной жидкости преобладают Na+ и С1–. Перемещение этих ионов через мембрану по специальным порам или каналам [50] в ответ на изменение электрохимического градиента создает трансмембранные токи [27, 32—35], способствующие возникновению потенциала действия. В состоянии покоя клеточная мембрана в принципе проницаема для К+ и относительно непроницаема для других внутри- и внеклеточных ионов [32]. Следовательно, разность потенциалов по обе стороны мембраны в покое определяется главным образом градиентом концентрации K+ [29, 31, 32, 48, 49]. Поддержание различий ионного состава и электрического заряда относительно мембраны в покое становится возможным благодаря двум факторам [31, 32, 49]: 1) свойствам проницаемости, или проводимости (g), клеточной мембраны для этих ионов, которые в свою очередь отражают способность мембранных каналов к их использованию теми или иными ионами; 2) функционированию различных ионных насосов и обменных механизмов [51—53], включая энергозависимый Na+/K+-обменный насос, перемещающий Na+ наружу, а К+ — внутрь клетки вопреки их градиентам концентрации [51, 53].
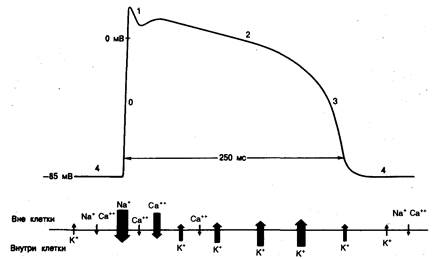
Рис. 4.2. Схематическое изображение потенциала действия в нормальном миокарде желудочков.
Показаны направление, величина и время протекания ионных токов, приводящих к развитию потенциала действия данной формы. Направление стрелки и ее размер указывают направленность и относительную величину тока (т. е. входящий или выходящий ток), а также соотношение амплитуд различных ионных токов. Горизонтальное положение стрелки соответствует моменту протекания токов по отношению к потенциалу действия. Обсуждение в тексте [36].
Потенциал действия: деполяризация. При стимуляции клетки и снижении мембранного потенциала до критического уровня (пороговый потенциал, при котором начинает генерироваться чисто входящий ток), происходит ряд последовательных изменений ионной проводимости мембраны и ионных токов, что обусловливает проявление потенциала действия [27, 31, 33, 36] (см. рис. 15.2). В норме деполяризация (фаза 0) полностью поляризованных сердечных волокон, за исключением клеток синусового и атриовентрикулярного узлов, обусловлена прежде всего резким увеличением мембранной проницаемости для Na+ (gNa) и быстрого входящего тока, переносимого ионами Na+ [24, 27, 29, 31, 33, 35, 48] (быстрый, направленный внутрь ток Na+, или быстрый входящий ток), в сочетании с открытием быстрых Nа+ каналов.
Способность мембраны к повышению gNa (т. е. к открытию закрытых Na+-каналов) зависит от уровня мембранного потенциала при возбуждении. Пропускная способность каналов для Nа+, как правило, максимальна, поэтому быстрый входящий ток Nа+ может иметь наибольшую величину, когда мембранный потенциал находится в пределах от — 85 до — 95 мВ. Если мембранный потенциал менее отрицателен, чем в норме, повышение проницаемости для Na+ и величина быстрого тока Na+ меньше максимально возможных из-за снижения пропускной способности каналов. При уменьшении мембранного потенциала ниже определенного уровня увеличения быстрого входящего тока Na+ может оказаться недостаточным для возникновения регенеративного или даже подпорогового ответа, что означает невозбудимость волокна.
Амплитуда и максимальная скорость деполяризации (Vmax) в нулевую фазу (нарастание) зависят от быстрого входящего тока Na+. Следовательно, они зависят и от уровня мембранного потенциала. Амплитуда сильно поляризованных волокон Пуркинье может достигать 130 мВ, a Vmax высока и, по некоторым данным, составляет от 500 до 1000 В/с. Обе величины уменьшаются при снижении уровня потенциала. На рис. 4.3, А показано постепенное уменьшение амплитуды и скорости нарастания потенциала действия волокна Пуркинье при снижении уровня мембранного потенциала. Кривые на рис. 4.3, Б показывают связь мембранного потенциала в момент возбуждения и Vmax ответа для двух клеток миокарда желудочков. Эту связь, впервые описанную для сердечных волокон Weidmann в 1951 г. [54] и позднее подтвержденную Hoffman и соавт. [55], часто называют «отношением реактивности», а кривую — «кривой реактивности». Такие кривые иногда используются для приблизительной оценки пропускной способности Na+-каналов.
Второй входящий ток в сердечной ткани [27, 29, 30, 33—35, 57, 58] активируется только при «низком» уровне мембранного потенциала — от — 35 до — 45 мВ. Этот ток переносится в основном ионами Са++ и имеет гораздо меньшую плотность (приблизительно 10%), чем быстрый ток Na+. Каналы, ответственные за его прохождение, активируются (открываются) и инактивируются (закрываются) медленнее, чем быстрые Na+-каналы, так что этот второй ток развивается медленнее и протекает дольше, чем быстрый ток Na+. Восстановление после инактивации также требует большего времени. Этот ток соответственно получил название «медленного входящего тока». Каналы, пропускающие быстрый и медленный входящий ток, являются фармакологически различными [58, 59]. Тетродотоксин избирательно блокирует быстрые каналы входящего Na+, тогда как медленные каналы блокируются такими агентами, как марганец (Mn++), D-600 и верапамил. В норме в сильно поляризованных клетках медленный ток оказывает лишь минимальное влияние на нарастание, внося основной вклад в фазу 2 (плато) реполяризации. С другой стороны, деполяризация узловых клеток с низким потенциалом [60—64], а также мышечных волокон створок атриовентрикулярных клапанов [66] и коронарного синуса [67], по-видимому, в большой степени определяется медленным током. Однако его вклад в деполяризацию неузловых волокон возрастает при снижении в них мембранного потенциала [30, 33—35, 66—69]. Этот последний факт приобретает особое значение, если учесть, что миокардиальные препараты, полученные у собаки после экспериментального инфаркта [70—74, 138, 184], а также у больных с органическим поражением сердца [21, 36, 76—90], содержат большое число частично деполяризованных волокон.
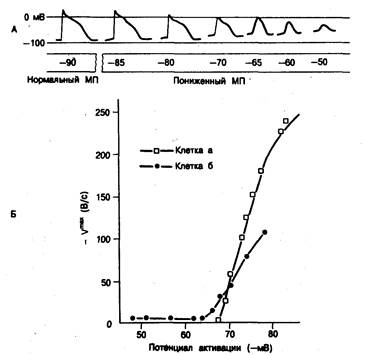
Рис. 4.3. Влияние потенциала покоя на потенциал действия.
А — изменения характеристик потенциала действия по мере его распространения из нормально поляризованной области в зону, где мембранный потенциал (МП) постепенно снижается. Величина МП (в милливольтах) указана для отдельных участков волокна; потенциалы действия, возникающие в этих точках, показаны выше. Следует, в частности, отметить постепенное уменьшение амплитуды и I^max, которое должно сопровождаться постепенным замедлением проведения. Обратите также внимание на временные изменения в ходе реполяризации и изменения длительности потенциала действия. Б — кривые, отражающие связь между уровнем мембранного потенциала возбуждения (в милливольтах по оси абсцисс) и (?щв«(в В/с по оси ординат) потенциала действия хорошо поляризованных клеток «а» (Еm = —90 мВ) и клеток «б» (Еm = —79 мВ) в волокнах желудочкового миокарда человека; образцы миокарда получены у больного с ишемической болезнью сердца и аневризмой желудочков. Измерения осуществлялись для потенциалов действия, возникающих при различных уровнях мембранного потенциала вследствие стимуляции препарата с определенными интервалами, во время реполяризации и фазы 4. Кривые, отражающие Эти переменные, называют «кривыми реактивности». Обратите внимание, что при средних значениях потенциала кривая для клетки «б» смещена вниз и вправо, а при более низких значениях — вверх и влево. Обсуждение в тексте.
Электрофизиологические свойства волокон, зависящих от медленного входящего тока (волокна с медленным ответом), отличаются во многих отношениях от свойств волокон, активность которых зависит от быстрого тока (волокна с быстрым ответом) [30, 33, 34, 58, 59]. Как правило, волокна с медленным ответом характеризуются низким диастолическим потенциалом, невысокой амплитудой и медленным нарастанием потенциалов действия, а также пониженной возбудимостью, измененной реактивностью, более продолжительной рефрактерностью и замедленным проведением. Кроме того, даже волокна, в норме не обладающие автоматизмом, могут приобрести способность к спонтанному генерированию импульсов при снижении мембранного потенциала до уровня, при котором начинает функционировать медленный ток (рис. 4.4, А). Большинство специфических черт активности синусового и атриовентрикулярного узлов удается объяснить, исходя из особенностей поведения медленного входящего тока [25, 30, 61—65]. То же справедливо для аномальных электрофизиологических характеристик многих частично деполяризованных клеток в ишемизированных и пораженных тканях сердца [31, 34, 36, 75, 78, 85, 86, 91—93].
Потенциал действия: реполяризация. Увеличение быстрого входящего тока Na+, которое предшествует деполяризации, является самоограниченным и непродолжительным (приблизительно несколько миллисекунд). Инактивация Na+-каналов с последующим уменьшением тока Nа+ дает начало реполяризации, процессу более длительному (до нескольких сотен миллисекунд) и сложному, в котором принимают участие ионы Na+, К+, Са++ и Cl– (см. рис. 4.2) [27, 29, 31, 33, 94]. Его специфические механизмы пока полностью не установлены. Ясно, однако, что для возникновения реполяризации необходимо уменьшение положительного заряда внутри клетки. В процессе реполяризации волокон Пуркинье можно выделить 3 четкие фазы. Начальная стадия (фаза 1) является быстрой; она обусловлена инактивацией тока Na+" и развитием выходящего тока, переносимого ионами К+ и, возможно, Сl– [27, 95]. После фазы 1 реполяризация существенно замедляется (плато, или фаза 2). В фазу плато происходит снижение суммарной проводимости мембраны и возникающие при этом токи небольшой величины более или менее уравновешивают друг друга [27, 33, 94, 95] (т. е. реполяризующие влияния инактивации тока Na+ и активации выходящего тока K+ уравновешиваются деполяризующим действием остаточного медленного входящего тока). Ток К+ со временем возрастает. Таким образом, вместе с уменьшением входящих токов это приводит к увеличению чистых потерь положительного заряда и, наконец, к быстрой реполяризации (фаза 3). По мере реполяризации калиевая проводимость (gK) все более повышается, способствуя дополнительному выходу К+ из клетки и дальнейшему ускорению реполяризации. Кроме того, процессу реполяризации способствует накопление вышедшего из клетки K+ в ограниченном внеклеточном пространстве [96]. Недавно была высказана гипотеза о том, что определенный вклад в реполяризацию может вносить электрогенный натриево-калиевый насос [97, 98]. Как только мембранный потенциал доходит до уровня примерно —40 или —45 мВ, начинается его быстрое снижение до значений потенциала покоя.
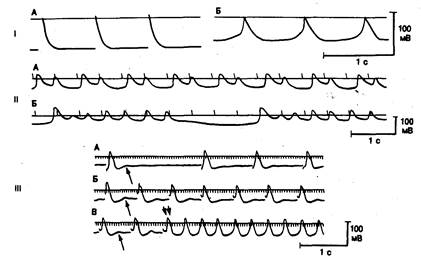
Рис. 4.4. Трансмембранные потенциалы в образцах миокарда предсердия (фрагменты I и III) и желудочка (фрагмент II) человека, позволяющие продемонстрировать различные типы аномального автоматизма в сердечной ткани при заболевании сердца.
Фрагмент I: трансмембранные потенциалы в хорошо поляризованном волокне с быстрым ответом (А) и низкоамплитудные медленные ответы частично деполяризованного волокна (Б) в придатке правого предсердия человека. Максимальный диастолический потенциал (и амплитуда), зарегистрированный в клетках А и Б, составили —72 мВ (—80 мВ) и —55 мВ (—55 мВ) соответственно. Высокоамплитудные, быстро нарастающие потенциалы, развивающиеся в сильно поляризованном волокне, существенно отличаются от низкоамплитудных, медленно нарастающих потенциалов в частично деполяризованном волокне. Следует также отметить, что клетка с медленным ответом начинает спонтанно деполяризоваться сразу после окончания реполяризации, т. е. она становится автоматически активной (автоматизм, вызванный деполяризацией).
Фрагмент II: трансмембранные потенциалы в частично деполяризованной, спонтанно активной клетке папиллярной мышцы больного с ревматическим заболеванием сердца, осложненным мерцанием предсердий, желудочковой эктопией высокой степени и вариабельными нарушениями желудочкового проведения, зависящими от частоты сердечного ритма. А — реполяризация после основного автоматического возбуждения прерывается однократной ранней постдеполяризацией, приводящей к развитию бигеминии. Б — каждое основное возбуждение прерывается низкоамплитудной осцилляцией, за которой следуют 5 повторяющихся высокоамплитудных ответов с результирующим выраженным увеличением длительности реполяризации. Каждая серия заканчивается колебанием мембранного потенциала очень низкой амплитуды, что еще больше затягивает окончание реполяризации при основном возбуждении. Фрагмент III: индукция колебаний мембранного потенциала по типу задержанной постдеполяризации и развитие триггерной активности в препарате предсердия человека, исходно не обладавшем спонтанной активностью. Первая осцилляция на каждой записи указана стрелкой. Записи А и Б, а также начало записи В демонстрируют развитие постдеполяризации при стимуляции с возрастающей частотой. Отмечается постепенное увеличение амплитуды осцилляций при повышении частоты стимуляции. В конце концов, колебания мембранного потенциала достигают порогового уровня и возникает ритмическая триггерная активность (в середине записи В двойная стрелка), которая сохраняется даже после прекращения стимуляции. Хотя каждый из приведенных примеров получен на частично деполяризованном препарате, аналогичные явления наблюдаются и в сильно поляризованных тканях. Обсуждение в тексте [87}.
Генерирование импульсов
Возбуждение и проявление потенциала действия обусловлены протеканием через клеточную мембрану достаточно деполяризующего тока, который быстро снижает (т. е. делает менее отрицательным) трансмембранный потенциал до порогового уровня. Для разных типов клеток пороговый потенциал различен и связан с максимальным диастолическим потенциалом [33, 56, 87, 99]. Возбуждающие (деполяризующие) токи могут поступать из внешнего источника или же развиваются спонтанно. В норме они являются следствием локальной разности потенциалов, возникающей при распространении потенциала действия. Определенные клетки, называемые автоматическими, способны генерировать такие токи спонтанно; следовательно, они могут испытывать самовозбуждение и спонтанно (автоматически) инициировать импульсы [25].
В норме автоматизм обусловлен периодическим возникновением в специализированных тканях сердца спонтанный деполяризации в фазу 4 [25] (см. рис. 4.1, Б). Клетки с наибольшей частотой спонтанной диастолической (фаза 4) деполяризации (в норме эти клетки синусового узла) служат первичными пейсмекерами, остальные же — представляют собой латентные пейсмекеры. В действительности деполяризация в фазу 4 развивается, скорее всего, только в клетках синусового узла (см. рис. 4.1, Б), в латентных же («ускользающих») водителях ритма необходимые изменения возникают лишь в ответ на такие факторы, как замедление синусового ритма или АВ-блокада.
Спонтанное генерирование импульсов может быть обусловлено не только медленной диастолической деполяризацией специализированных автоматических клеток, но и другими механизмами. Поскольку такие альтернативные механизмы являются недостаточными для работы в нормальных физиологических условиях, их рассматривают как аномальные факторы автоматизма, а возникающую в результате спонтанную активность — как аномальный автоматизм [100]. Этот термин объединяет целую группу различных явлений. Спонтанные циклические пейсмекероподобные осцилляции диастолического потенциала, не связанные с основным инициирующим событием, представляют один из важнейших типов активности. Такого рода активность проявляется преимущественно в угнетенных, частично деполяризованных волокнах, где мембранный потенциал снижен до уровня, при котором начинает функционировать медленный входящий ток; отсюда название «автоматизм», вызванный деполяризацией». По-видимому, подобная активность может возникать при снижении диастолического потенциала в большинстве клеток (если не во всех), включая рабочий миокард предсердий и желудочков [101—103]. Такое явление часто наблюдается в ишемизированных и пораженных тканях сердца [36, 74, 75, 87, 91—93]. Вполне репрезентативна в этом отношении запись на рис. 4.4 (фрагмент 1), полученная в частично деполяризованных автоматических клетках миокарда предсердий человека.
Существуют и другие типы осцилляций, которые зависят от предшествующего инициирующего потенциала действия (т. е. триггерная активность). Такие осцилляции могут возникать во время (или после завершения) реполяризации инициирующего потенциала действия (см. рис. 4.4, фрагменты II и III). Для их обозначения был предложен целый ряд определений, из которых наиболее широко используются термины, предложенные Cranefield: соответственно «ранняя» и «задержанная» постдеполяризация [30]. Ранняя постдеполяризация чаще всего прерывает реполяризацию в фазу 2 на плато при уровне потенциала от —20 до —25 мВ, но она может возникнуть и во время фазы 3. Она увеличивает длительность реполяризации и рефрактерности иногда на многие секунды (см. рис. 4.4,11). Задержанная же деполяризация (см. рис. 4.4, III) наблюдается во время фазы 4 при более или менее нормальном уровне диастолического потенциала (от —70 до —185 мВ), а также при его сниженном уровне. Постдеполяризация, достигающая порогового уровня, приводит к возникновению триггерных ритмов (см. рис. 4.4,III) [104], из которых лучше всего известен ритм, обусловленный интоксикацией сердечными гликозидами.
Мембранные механизмы, лежащие в основе возникновения пейсмекерных потенциалов, до конца не выяснены [29, 30, 33]. Согласно последним, работам, пейсмекерная активность волокон Пуркинье является следствием активации входящего тока, переносимого как Na+ так и K+ [105, 106]. По некоторым данным, в синусовом узле определенный вклад в пейсмекерную активность вносит и медленный входящий ток [107—НО]. Ионные механизмы, определяющие автоматическую активность частично деполяризованных миокардиальных клеток и осцилляции постдеполяризационного типа, еще менее изучены [29, 30, 33, 34, 36, 102, 103].
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 461; Нарушение авторских прав?; Мы поможем в написании вашей работы!