КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Дегенерація олігодендроцитів та їх відновлення після фокальної ішемії мозку
|
|
|
|
Вперше вивченням проблеми загибелі олігодендроглії за умов фокальної ішемії та розробкою методів відновлення її клітин зайнялися команда американських дослідників з Washington University School of Medicine, очолювана професором Саллі МайАйвер (Sally R. McIver). Дослідження проводилися з використанням дорослих щурів-самців лінії Sprague Dawley вагою 200-250 г (Charles Rivers Laboratories, Wilmington, Massachusetts) [50] (рис. 2. 11).

Рис. 2. 11. Щури лінії Sprague Dawley
При виконанні роботи авторами були використані наступні терміни та позначення: BrdU – бромодеоксиуридин, CBF – мозковий кровотік, eGFP – enhanced green fluorescent protein (зелений флуоресцентний білок), LFB – барвник Luxol Fast Blue, LV – лентівірусний вектор, MBP (ОБМ) – основний білок мієліну, MCAО – оклюзія серединної мозкової артерії, OPC – клітини-попередники олігодендроцитів, SVZ – субвентрикулярна зона мозку, TTC – трифеніл тетразоліум хлорид, TUNEL – трансфераза термінального приєднання дезоксиуридинтрифосфату (маркер загиблих олігодендроцитів).
Вченими показано, що вразливість олігодендроцитів за умов церебральної ішемії сприяє втраті функціонально повноцінних мієлінових оболонок. Це може призводити до руйнування білої речовини мозку. Сучасні іммуноцитохімічні методи виявлення ушкоджень олігодендроцитів в експериментальних моделях базуються на використанні антитіл до специфічних епітопів, наявних у структурі ОБМ. При цьому ці методи не дискримінують структурних змін в олігодендроцитарній морфології [50].
В попередніх роботах дослідники описували використання лентівірусного вектору (LV) (рис. 2. 12), що несе сенсибілізований ген еGFP, розташований поряд з промотором гену ОБМ [51]. Це використано для селективної візуалізації ушкоджених ішемією, а також дозволяє оцінити ступінь ураження тканини мозку. У цьому дослідженні [50] вченими використано LV-MBP-еGFP для визначення олігодендроцитів у білій речовині головного мозку щурів за умов перехідної фокальної церебральної ішемії та встановлено ступінь ушкодження олігодендроглії через 24 годин, 48 годин і один тиждень після реперфузії шляхом оцінки кількісного виживання клітин і аналізу процесів мієлінізації. Вірусний вектор LV проникав у клітини олігодендроглії та вбудовувався в їх геном. При цьому, як було зазначено вище, вектор містив у своєму складі сенсибілізований еGFP та ген ОБМ [50].
Рис. 2. 12. Структура капсиду лентівірусу
В загиблих у результаті церебральної ішемії олігодендроцитах спостерігається невисока реплікативна активність вірусу, причому було відмічено прогресуючу втрату GFP(+)-клітин через 24 та 48 годин після індукції ішемічного ураження мозку. GFP(+)-клітини, які вижили, мали змінену морфологію ядра, яка нагадувала аналогічну морфологію пікнотичного ядра при некрозі. Проте їх ядро не піддавалося некротичним ураженням. Такі клітини не мали у своєму складі ферменту TUNEL, тобто були TUNEL-негативними. У таких клітин показано порушення процесів мієлінізації, в результаті чого фрагментована мієлінова оболонка на початку 24-ї години після ішемії не відновлювалася.
Через тиждень після ішемії, дослідники спостерігали відновлення популяцій GFP(+)-олігодендроцитів, що було добрим прогностичним показником при відновленні мієліногенезу. Проте, за допомогою BrdU- включень показано, що проліферуючі клітини-попередники олігодендроглії не були основним джерелом GFP(+)-олігодендроцитів. Ці спостереження ідентифікували наявність нових перехідних клітинних форм між попередниками і зрілими олігодендроцитами. Подальше вивчення таких нових попередників дозволить відкрити нез'ясовані механізми структурних змін мієліну при ішемічній патології мозку [50].
Виживання і збереження цілісності мієлінопродукуючих олігодендроцитів має вирішальне значення для нормальної функції аксонів білої речовини мозку. Існує все більше доказів на моделях хвороби і в трансгенних нокаут-мишей, позбавлених експресії білків мієліну, що первинна дисфункція в олігодендроцитах може призвести до вторинного пошкодження аксонів. Це спостерігається при різноманітних неврологічних порушеннях, зокрема розсіяному склерозі, інсульті, перинатальній травмі головного мозку тощо [52].
Уразливість олігодендроцитів за умов ішемічної пошкодження мозку продемонстрована в численних моделях в пробірці і опосередковується як окисними, так і ексайтотоксичними механізмами. При гострій ішемії, яка розвинулася в результаті травматизації головного чи спинного мозку продемонстровано втрату імунореактивності до олігодендроцит-специфічних маркерів і появу у клітинах пікнотичних ядер. Олігодендроцит-специфічна експресія флуоресцентних маркерів в умовах ішемічного інсульту демонструє зниження інтенсивності флуоресценції в мієліновій оболонці.
Дані ішемічного ушкодження білої речовини в природних умовах показані на ранніх тимчасових точках при фокальній ішемії, коли лише спостерігаються невеликі ультраструктурні змін в морфології мієлінових оболонок. Гістологічні та імуноцитологічні методи, використані у постійних і перехідних координаційних моделях ішемії вказали на відносну втрату імунореактивності для олігодендроцитів і білків мієліну, починаючи вже з 24 години після індукції ішемії [53].
У пробіркових моделях ішемія піддається фармакологічним маніпуляціям. Такі моделі є цінними для вивчення гострої відповіді олігодендроцитів при пошкодженні мозкової тканини, однак є неповним відображення того, що відбувається в природних умовах, де судинні і запальні реакції є додатковими факторами ушкодженння мієліну. У природних умовах дослідження були обмежені наявністю епітопів в пошкодженій тканині та забезпечили дослідників інформацією відносно невеликих структурних змін, які можуть відбуватися різними шляхами в сомі олігодендроцитів. Структурні зміни в олігодендроциті, що відбуваються у відповідь на ішемію, добре визначені при проведенні тривалих досліджень.
Зміна перебігу процесів ремієлінізації може сприяти функціональному відновленню при демієлінізуючих захворювань і може бути залученою в початкове відновлення функції клітин в моделі пошкодження спинного мозку. Як стверджують деякі дослідники, цього можна досягти, якщо замість ушкоджених клітин трансплантаційним шляхом ввести нові, генномодифіковані попередники олігодендроглії. Тим не менш, не цілком сформований потенціал для подібної заміни олігодендроцитів при ішемії білої речовини мозку, особливо якщо це стосується дорослого організму [50].
У мозку новонароджених тварин є запас недиференційованих клітин, які за умов травматизації мозку під дією ендогенних сигнальних молекул можуть утворювати різні класи клітин нервової системи. Так може утворюватися клас специфічних «білих» олігодендроцитів, які є достаньо стійкими при різних патологічних станах мозку. Проте для дорослого мозку така кількість клітин є набагато меншою, тому питання про формування «особливих» резистентних олігодендроцитів залишається відкритим. Визначення долі «білих» олигодендроцитів речовини після осередкової ішемії в довгострокових дослідженнях може дати уявлення про передбачувані ендогенні механізми репарації мієліну, такі як ті, що описані при у моделях травматичного пошкодження спинного мозку.
Саллі МакАйвер так коротко описує схему проведення досліджень. Рекомбінантний лентівірус створено на основі вірусу імунодефіциту людини. До складу рекомбінанту внесено невеликий 1,9 кб (кб = 1 кілобаза = 1000 пар нуклеотидів у ДНК) фрагмент промотора ОБМ разом з геном GFP. Рекомбінантний лентівірус був підготовлений перехідною ко-трансфекцією клітинної лінії НЕК 293Т з вектором МВР-EGFP та лентивірусів допоміжних плазмід (pMDLgpRRE, RSV-REV і PMDG для VSV-G pseudotyping) з використанням методу соосадження фосфатом кальцію. Обмін генетичним матеріалом міжвірусними геномами здійснився через 24 год і через 48 год трансфекції дослідники збирали лентівірусів, фільтрували через фільтр 0,45 мкм і концентрували за допомогою ультрацентрифугування (90 хвилин при 25000 обертів на хвилину), а потім ресуспендували в розчині Рінгера. Зараження клітин НЕК 293 з вірусом був використане для титрування за допомогою проточної цитометрії через 24 години після інфікування. Аліквоти вірусних суспензій зберігали при -80 ° С до використання [50].
Дорослих Sprague Dawley самців щурів (Charles Rivers Laboratories, Вілмінгтон, штат Массачусетс) з масою тіла 200-250 г використовували в цьому дослідженні. Хірургія тварин і догляд проводилися відповідно до «Керівництва по догляду та використання лабораторних тварин» Інституту науково-дослідної лабораторі (ILAR). Всі процедури були схвалені Комітетом тваринних досліджень при Вашингтонському університеті.
Щурів анестезувати ізофлураном (3% індукції, 1,75% з технічного обслуговування (Stoelting, Wood Dale, Іллінойс). Після анестезії робили серединний розріз голови, при цьому шприць Гамільтона опускали в ліву бокову частину мозолистого тіла через невеликий отвір. Комбінований вектор LV-МВР-EGFP (4,5 мкл) був доставлений зі швидкістю 0,45 мкл / хв через наноін'єктор насоса (Stoelting), після чого голка шприця була залишена на місці протягом 5 хвилин, щоб забезпечити повне поширення вірусу [50].
Через шість днів після ін'єкції, щури піддавалися або фіктивній хірургії (Sham-контрольні тварини), або впливу перехідної фокальної ішемії за допомогою 60-хвилинної оклюзії середньої мозкової артерії (МСАО) внутрипросвітньою нейлоновою ниткою. Розрізи зашивали і тваринам давали відновитися після анестезії під час оклюзії. Після 60 хвилин MCAO щурів повторно анестезувати та припиняли ішемію шляхом видалення внутрішньопросвітнього шва. Дослідники використовували лазерний доплерівський зонд для вимірювання мозкового кровотоку (CBF) в різні моменти часу, щоб підтвердити наявність ішемічних станів тканини мозку: CBF вимірювали безпосередньо над територією MCA на початку оклюзії і до реперфузії, щоб підтвердити стійкий зниження кровотоку (<20 % вихідного CBF), і на початку реперфузії, щоб підтвердити достатнє відновлення кровотоку (> 60% CBF). Тварини, що не відповідають цим критеріям, були виключені з дослідження. Sham-контрольні тварини були піддані тим же процедурам, без оклюзії МСА внутрішньопросвітним швом.
Тварини були розділені на наступні групи, відповідно до MCAO: 24 години реперфузії (n = 11 досліджуваних, 7 Sham-контрольних); 48 годин реперфузії (n = 11 досліджуваних, 7 Sham-контрольних); 1 тиждень реперфузії (n = 12 з ішемічним інсультом, 12 Sham-контрольних). Смертність, як правило, від затримки неповної реперфузії і / або кровотечі, був рівною серед груп (n = 2-3 на групу). Щоб відстежувати долю проліферуючих клітин, окремий набір тварин (N = 4-х тактний, 1 Sham-контроль) щодня отримував внутрішньочеревну ін'єкцію 50 мг/кг BrdU (Sigma-Aldrich, Сент-Луїс, штат Міссурі) в день після інсульту. Тварин умертвляли через один тиждень після реперфузії [50].
Щурів умертвляли в кожному певному пункті часу внутрішньосерцевою перфузії з PBS (potasium buffer solution, забуферений фосфатний розчин). Мізки тварин були витягнуті з наступним поділом на зрізи товщиною 3 мм. Зрізи інкубували у розчині хлориду трифеніл тетразолію (ТТС; Sigma-Aldrich) протягом 3 хвилин при 37 ° С, фіксували в 4%-му р-ні параформальдегіду протягом 24 годин, і піддавали кріопротекціі через градієнти сахарози (10-30%) протягом принаймні 24 годин кожен. TTC-пофарбовані зрізи були відскановані для документації розміру інфаркту та 20-мікрометрові послідовні кріозрізи були зібрані (кожні 4 частини) на 3 набори слайдів. Один набір був використаний для швидкого фарбування барвником LFB наступним чином: кріозрізи були зневоднені в 70, 80 і 95%-му етанолі (EtOH, 5 хв кожен), інкубовані в 1% LFB (в 95%-му EtOH + 10%-ої оцтової кислоти) протягом ночі при 60°С в герметичному контейнері, промивають у 95%-му EtOH, водою (2 хв), 0,05%-м карбонатом літію (1 хв), 70%-м етанолом і водою. Останні три кроки повторюються до тих пір, поки не отримають оптимальний контраст між білою і сірою областями речовини мозку.
Другий набір кріозрізів був використаний кількісної і морфологічної оцінки GFP(+)-олігодендроцитів. Третій сет був використаний для імуногістохімічного виявлення олігодендроцитів клітин-попередників (OPCs; NG2) або колокалізаціі з маркерами клітинної проліферації (BrdU) або загибелі (TUNEL). Зрізи промивали PBS (3 рази кожні 5 хв) і блокували протягом 30 хвилин з використанням 2% нормальної козячої сироватки (Sigma) і 0,1%-го Тритон-X (Sigma) в PBS. Тканину інкубували протягом ночі при 4°С в анти-кролячій NG2 (Millipore, Billerica, Massachusetts; 1: 100) або анти-BrdU блокуючій сироватці кролика (1: 1500; Мегабаза, Нью-Йорк); потім промивали розчином PBS (3 рази кожні 5 хв), та інкубували протягом 1-2 годин при кімнатній температурі з козячими анти-кролячими Alexa-594 вторинними антитілами (Invitrogen; 1: 1000). Виявлення BrdU проводили з 10-хвилинною інкубацією в 2 н. HCl при 37°С з подальшою 20-хвилинною інкубацією в розчині 0,2 М борної кислоти, до стадії блокування процесу.
Для виявлення загибелі клітин маркер TUNEL (Millipore) був використаний відповідно до протоколу. Всі зрізи обробляли з Hoechst 33342 методом накладення ядерних плям (Invitrogen) та досліджували за допомогою лазерної скануючої конфокальної (Zeiss, Maple Grove, Міннесота) мікроскопії. Цифрові кольорові мікрофотографії були додатково оброблені за допомогою програми Adobe Photoshop [50, 51].
Розуміння механізмів ішемічного ушкодження в природних умовах в моделі ішемії є необхідним кроком на шляху до розробки терапевтичних втручань. У фокальній ішемії людини, інфаркти часто пов'язані з ураженням білої речовини, і ретельне вивчення клітинного пошкодження в білій речовині має вирішальне значення в моделях інсульту тварин. Попередні дослідження вивчали відносну зміні експресії олігодендроцит- і мієліноспецифічних маркерів, але ці епітопи можуть бути тимчасово втрачені, або замасковані при патологічних станах. Навіть у здорової тканини, візуалізація антитіл до цих антигенів, як правило, обмежена або в тілі клітини або товстим шаром мієліну, і може не показувати всіх змін у морфології олігодендроцитів, які відбуваються у відповідь на пошкодження. Трансгенні миші, які продукують олігодендроцитарні клони клітин з конкретними флуоресцентними мітками, дозволяють повністю візуалізувати мофологію олігодендроцитів, і надають розуміння ішемічного ушкодження цих клітин. Тим не менш, в лініях трансгенних тварин, отриманих нині, візуалізація флуоресцентних міток може бути вираженою в клітинах-попередниках, а не лише зрілих олігодендроцитах. Крім того, високі рівні експресії в трактах білої речовини ускладнюють розмежування окремих процесів мієліногенезу, і, отже, обмежують вимірювання аналізів, які нечутливі до тонких змін у морфології.
Електронна мікроскопія (ЕМ) має високу чутливість до виявлення клітинного пошкодження і використовується для опису ультраструктурних змін в олігодендроцітах, що відбуваються в ранні терміни після ішемії. EM забезпечує морфологічний деталі, необхідні для оцінки тонких відмінностей в життєздатності клітин, але для ультраструктурного кількісного аналізу травматизованої зони слід здійснювати повторний відбір біозразків в селективних, невеликих регіонах, які нерідко достатньо складно виявити. Крім того, більшість цих досліджень були обмежені виміром різких змін при ішемії білої речовини мозку [53].
Мало що відомо про часову залежність дегенеративних змін або регенеративного потенціалу білої речовини від тривалості дії фактору індукції ішемічного ураження. У цьому дослідженні вчені використовували лентивірусний вектор спеціально для зрілої мієлінпродукуючої популяції олігодендроцитів щурів в білій речовині. Направляючи LV-MBP-EGFP ін'єкцією у бічне мозолисте тіло, дослідники отримали зону поширення проліферуючих клітин при реперфузії. Конфокальна мікроскопія застосовувалася для того, щоб більш точно дослідити часовий хід відновлення білої речовини після MCAO. Використання TTC та фарбування LFB підтвердили, що ця модель ішемії провокує виникнення інфарктів, пов'язаних з ушкодженням білої речовини, як повідомлялося раніше. TTC-забарвлені тканини використовували для документування загальної площі інфаркту, а потім згодом обробляли для візуалізації травми білої речовини на клітинному рівні. Це важливо, оскільки MCAO може індукувати появу різних за розміром інфарктів, навіть якщо вимірювання потоку крові головного мозку послідовно знижується під час оклюзії. Тут використання ТТС дозволяє вченим стверджувати, що зменшення кількості GFP(+)-олігодендроцитів в бічному мозолистому тілі починається вже з 24-48 годин реперфузії після інсульту. Щодо розподілу GFP(+)-олігодендроцитів, які спостерігаються в білій речовини, дослідники також спостерігали залежне від часу зменшення числа тих клітин, що уздовж медіально-бічної зони мозолистого тіла, при майже повній відсутності в зовнішній зоні через 48 годин після ішемії [50].
Малоймовірно, що зниження експресії GFP відбувалося через неактивність промотора MBP, оскільки рівень транскрипції MBP збільшується після розвитку ішемії. GFP(+)-олігодендроціти, що вижили, були виявлені в регіонах ішемічної загибелі клітин, які мали у своєму складі маркер TUNEL. Враховуючи ці дані, то малоймовірно, що зниження GFP(+)-клітин пригнічується активністю промотора MBP.
Широкий некроз в смугастому тілі (corpus striatum) і корі є визначальною рисою серцевини інфаркту в цій моделі ішемії, тому цілком імовірно, що GFP(+)-олігодендроцити піддаються клітинній загибелі, що призводить до втрати або деградації GFP. Тут вчені прагнули вивчити олігодендроцити ближче до межі ішемізованої зони, де може бути більший потенціал клітин для відновлення. Тому що варіабельність в розмірах інфаркту є основною рисою цієї експериментальної моделі, дослідники не могли виключити можливість того, що деякі клітини, проаналізовані в даному дослідженн, загинули саме в центральній зоні (ядрі) інфаркту [50].
GFP(+)-олігодедроцити, що вижили, характеризувалися зниженням мієліноутворюючого потенціалу. При цьому були виявлені значні зони фрагментації мієлінових мембран на 24- і 48-у години після реперфузії. Живі GFP(+)-олігодедроцити характеризувалися також TUNEL(-)-показником та не мали пікнотичних ядер. Ці результати показують, що дегенерація процесів мієлінізації (мієліногенезу) може передувати загибелі олігодендроцитів [50]. Це переконливі перспективи у світлі доказів того, що на цей процес індукують високі концентрації глутамату. Показано, що за умов ішемії експресія субодиниць глутаматних рецепторів спостерігається саме в зонах демієлінізації [54].
Збереження морфології тіла клітини під час міелінової дегенерації представляє інтригуючу можливість того, що реміелінізація відбувається завдяки структурному відновленню раніше пошкоджених олигодендроцитів. На перший тиждень реперфузії після осередкової ішемії, спостерігалося відновлення інтактних процесів мієлінізації і кількості GFP(+)-олігодендроцитів в ішемізованій області білої речовини. Проте, як було зазначено раніше, порушення мієліногенезу спостерігалося через 24-48 годин після ішемії.
На додаток до відновлення структури олігодендроцитів спостерігалося і відновлення GFP(+)-клітин після ішемії білої речовини мозолистого тіла (corpus callosum). Репопуляціі GFP(+)-олігодендроцитів, візуалізовані в цьому дослідженні, швидше за все, походять від клітин-резидентів вже в білій речовині, а не клітин, що мігрують в районах з сірою речовиною.
Використання BrdU-реєстрації дозволило відстежити реакцію проліферуючих клітин протягом одного тижня після реперфузії, щоб визначити, диференціюються вони в GFP(+)-олігодендроцити, чи ні. Вчені знайшли численні BrdU-клітини в тій же області ішемічного ураження білої речовини, де були знайдені регіони збільшених популяцій GFP(+)-олігодендроцитів [50] (рис. 2. 13).

Рис. 2. 13. Локалізація BrdU- та GFP-позитивних клітин у мозолистому тілі мозку: А – зображення латеральної частини мозолистого тіла (ядра BrdU-позитивних клітин червоні, GFP-позитивні клітини – зелені); В і С – напрямки міграції BrdU(+)- та GFP(+)-клітин
З багатьох GFP(+)-олігодендроцитів менше 5% були BrdU-позитивних. Крім того, спільна локалізація з маркером олігодендроцитів, CC-1, показала, що тільки 11% олігодендроцитів, що знайдені в ішемізованій зоні білої речовини, мічені BrdU. Враховуючи ці дані, то малоймовірно, що молоді проліферуючі клітини відповідають повною мірою за відновлення зрілих олігодендроцитів. Можливо, що дані професора МайАйвер недооцінюють кількість GFP(+)-олігодендроцитів, отриманих з популяцій проліферуючих клітин. Тим не менш, / СС-1(+)-олігодендроціти BrdU було виявлено протягом одного тижня після травми спинного мозку з використанням аналогічного режиму дозування. Навіть одного імпульсу BrdU, враховуючи 24 годин після травматизації спинного мозку, було достатньо, щоб відстежувати новостворені олігодендроцити через тиждень [50, 53].
В даний час широко визнається, що дорослі OPCs несуть відповідальність за ремієлінізацію аксонів після ішемічного ушкодження. У моделях демієлінізації було добре показано, що OPCs реагують на втрату олігодендроцитів проліферацією і диференціацією в мієілноутворюючі олигодендроцити (рис. 2. 14). Наприклад, в спинномозкових моделях контузії втрата олігодендроцитів спостерігалася одночасно з підвищенням експресії NG2 і проліферації клітин в ранні терміни після травми, і йшла за рахунок збільшення олігодендроцитів протягом декількох тижнів після травми. У цих дослідженнях, заміна олігодендроцитів чітко простежувалася в сірих і білих областях мозку, що межують з ішемізованою ділянкою, і була пов'язана з функціональним відновленням. Крім того, в моделі демієлінізації, ретровірусні маркування ендогенно проліферуючих клітин показали, що NG2 (+)-попередники сприяли ремієлінізації. Мало уваги приділялося відповіді NG2(+)-клітин-попередників і заміні ними олігодендроцитів після ішемічного ушкодження в мозку дорослого організму, а існуючі дослідження були в основному обмежені розглядом клітинних змін в сірій речовині, де мієлінізація є мінімальною порівняно з білою речовиною. Тим не менше, ці дослідження показали, що залежність часового збільшення NG2-імунореактивності збігається з відновленням олігодендроцитів і мієліну в прилеглих до ішемії зонах мозку на 1-2 тижні реперфузії. У ядрі ішемії, однак, спостерігалася уповільнена втрата NG2 + OPCs, олігодендроцитів і мієліну [50].
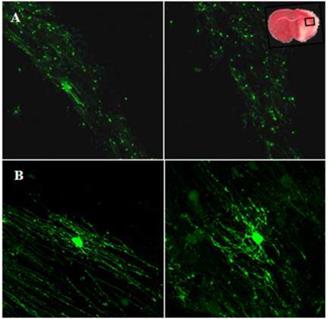
Рис. 2. 14. Стан олігодендроглії в ішемізованій зоні мозку: А – на 24 год після розвитку ішемії, В – через 1 тиждень після ішемії (клітина-попередник OPC дає початок зрілому олігодендроциту). На рис. зверху справа показано уражену ішемією ділянку мозку
Дослідники виявили збільшення NG2-імунореактивності всієї ішемізованої білої речовини, яке зберігалося на 1 тиждень реперфузії. В 48 годин після реперфузії, NG2(+)-клітини виявлялися гіпертрофічними, з високим ступенем мітотичної активності, однак зміненою морфологією. Через 1 тиждень після реперфузії, NG2 клітини відновлювали нормальну морфологію та індукували відновлення GFP(+)-олігодендроцитів. Враховуючи ці дані, цілком імовірно, що більшість відновлених GFP(+)-клітин, розглянутих у даному дослідженні, були отримані не від OPCs, які були інфіковані LV-MBP-EGFP [50].
У моделях травми спинного мозку і демієлінізації, джерела заміни клітин і визначальних чинників проліферації і диференціювання можуть відрізнятися від аналогічних у білій речовині головного мозку. Наприклад, в головному мозку субвентрикулярна зона (SVZ) є локальним джерелом активно проліферуючих клітин нервової ситеми [55, 56] (рис. 2. 15).

Рис. 2. 15. Зріз мозку ембріона щура. У бокових шлуночках головного мозку темною лінією позначено субвентрикулярну зону (SVZ), де локалізовані ізоформи ферменту глутаматдекарбоксилази 67 (GAD 67)
Нервові стовбурові клітини в SVZ реагують на ішемічне пошкодження за рахунок збільшення швидкості проліферації і зменшення довжини клітинного циклу. Крім того, МайАйвер виявила збільшення числа BrdU-клітин в SVZ в мозку інтактних тварин, у порівнянні з Sham-контрольними тваринами. Оскільки існує багато доказів того, що клітини субвентрикулярної зони сприяють нейрогенезу в прилеглих до ішемії областях кори шляхом міграції через мозолисте тіло, цілком можливо, що деякі з BrdU-клітин, які вчені знайшли в ішемізованій зоні білої речовини, були нейрогенними [56].
Клітини мікроглії також знайдені у межах цієї зони та смугастому тілі і може грати допоміжну роль в нейрогенезі. Хоча команда МакАйвер неуважно вивчила поширення мікроглії при ішемічному ураженні білої речовини, проте у дослідників спостерігалося збільшення імунореактивності для маркеру мікроглії, Iba-1 (рис. 2. 16). Ємність SVZ-клітин, отриманих для генерації олігодендроцитів, продемонстрована в демієлінізуючих ураженнях мозолистого тіла [55].
Рис. 2. 16. Структура білка Iba-1 (ionized calcium-binding adapter molecule 1) – головного цитозольного маркеру мікроглії
Відновлення функції білої речовини зазвичай відбувається у випадках з людським інсультом, а в тваринних моделях інсульту він пов'язаний з проростанням аксонів. Це буде дуже корисно при з'ясуванні ендогенних шляхів, що ведуть до відновлення олігодендроцитів після ішемічної демієлінізації, так як ці механізми можуть також сприяти відновленню функцій білої речовини після інсульту [50].
Команда МакАйвер та інші дослідники наводять докази, що підтверджують здатність до регенерації білої речовини, яке залежить від кількості відновлених клітин і збереження процесів мієліногенезу. Лентівірусні вектори не тільки підвищують візуалізацію морфологічних змін олігодендроцитів у відповідь на пошкодження, а й забезпечують ефективний метод для оцінки потенційних захисних ефектів нової трансгенної експресії. Це може бути корисним підходом для діагностики багатьох захворювань, при яких активація процесів ремієлінізації ушкоджених після травми ділянок мозку є терапевтичною метою.
ВИСНОВКИ
На основі розглянутого у роботі літературного огляду можна зробити наступні висновки:
1) Мієлінові мембрани – обов'язкова складова нервової тканини, ключовою функцією яких є інтеграція передачі нервового збудження. Обмін білково-ліпідних компонентів мієліну постійно змінюється протягом онтогенезу.
2) Структура мієлінової оболонки може порушуватися за умов різних патологічних станів нервової системи або всього організму в цілому. При руйнуванні мієлінової оболонки відбувається накопичення у тканинах мозку основного білка мієліну (ОБМ). Між концентрацією ОБМ і ступенем деструкції (дезагрегації) мієліну існує пряма залежність.
4) За умов ішемічного ураження білої речовини мозку спостерігаюється суттєві зміни властивостей мієлінової оболонки. HIF-1α у високих концентраціях може блокувати життєдіяльність олігодендроглії, що, у свою чергу, припиняє мієліногенез. Специфічні пептидази та сфінгомієлінази в цих умовах руйнують мієліновий каркас, тим самим порушуючи проведення нервового імпульсу.
5) Деякі нейротропні препарати (енцефабол, пікрозид, α-кетоглутарат тощо) позитивно впливають на відновлення олігодендроглії, модулюючи мієліногенез (мієлінізацію).
6) Використання вірусних векторів, зокрема, лентівірусних рекомбінантів, дозволяє підвищити візуалізацію морфологічних змін олігодендроцитів у відповідь на ішемічне ушкодження, і забезпечити ефективний метод для діагностики нейродегенеративних захворювань нервової системи, наприклад, ішемічного інсульту.
|
|
|
|
|
Дата добавления: 2015-06-27; Просмотров: 718; Нарушение авторских прав?; Мы поможем в написании вашей работы!