КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Биомеханика мышцы
|
|
|
|
Поперечно-полосатая структура мышечных волокон может наблюдаться под обычным микроскопом. Отдельное мышечное волокно имеет диаметр 20 - 80 мкм и окружено плазматической мембраной толщиной 10 нм. Каждое отдельное волокно - это сильно вытянутая клетка. Длина отдельных волокон {клеток) может существенно варьироваться в зависимости от вида мышцы от сотен микрон до нескольких сантиметров. Внутри волокна, кроме известных органелл (ядро, ядрышко, митохондрии, аппарат Гольджи и др.), находятся сократительный аппарат клетки, состоящий из 1000 - 2000 параллельно расположенных миофибрилл диаметром 1-2 мкм, а также клеточные органеллы: саркоплазматический ретикулум и система поперечных трубочек - Т-система.
Структура поперечно-полосатой мышцы. Модель скользящих нитей.
БИОФИЗИКА МЫШЕЧНОГО СОКРАЩЕНИЯ.
Регистрация и анализ данных функциональных исследований
Приборы для измерения функции внешнего дыхания. Устройство и принцип работы.
Составители: Крылова Л.А.
Ковалёва Л.В.
Мышечная активность – это одно из общих свойств высоко организованных живых организмов. Вся жизнедеятелъность человека связана с мышечной активностью. Независимо от назначения, особенностей строения и способов регуляции принцип работы различных мышц организма одинаков.
Мышечная клетка отличается от других возбудимых клеток таким специфическим свойством, как сократимость, т.е. способность генерировать механическое напряжение и укорачиваться. Кроме того, мышцы являются генератором тепла, причем не только при мышечной работе, холодовой дрожи, но и в режиме термогенеза.
Мышечная активность о процессе жизнедеятельности обеспечивает работы отдельных органов и целых систем: работу опорно-двигательного аппарата, легких, сосудистую активностъ, работу желудочно-кишечного тракта, сократительную способность сердца. Нарушение работы мышц (например, определяющих функционирование легких, сердца) может приводить к патологиям, а ее прекращение - даже к летальному исходу.
Мышечная ткань представляет собой совокупность мышечных клеток (волокон), внеклеточного вещества (коллаген, эластина и др.) и густой сети нервных волокон и кровеносных сосудов. Мышцы по строению делятся на: гладкие - мышцы кишечника, стенки сосудов, и поперечно-полосатые — скелетные, мышцы сердца. Независимо от строения все они имеют близкие механические свойства, одинаковый механизм активации и
близкий химический состав.

Справа – схема расположения актина и миозина на поперечном срезе.
В миофибриллах (Рис.1) различают: А -зону - темные полосы, которые в поляризованном свете дают двойное лучепреломление, т.е. обладают свойством анизотропии (отсюда и название: А -зона), I -зону - светлые полосы, не дающие двойного лучепреломления, то есть изотропные (отсюда название: I -зона). В области I-зоны проходит темная узкая полоса –
Z-диск. Промежуток между двумя Z-дисками называется саркомером и является элементарной сократительной единицей мышечной клетки.
Саркомер - это упорядоченная система толстых и тонких нитей, расположенных гексагопально в поперечном сечении. Толстая нить имеет толщину - 12 нм и длину - 1,5 мкм и состоит из белка миозина. Тонкая нить имеет диаметр 8 нм, длину 1 мкм и состоит из белка актина, прикрепленного одним концом к Z-диску.

Рис.2. Микроструктура саркомера.
Актиновая нить состоит из двух закрученных один вокруг другого мономеров актина толщиной по 5 нм (рис. 2). Эта структура похожа на две нитки бус, скрученные по 14 бусин в витке. В цепях актина регулярно примерно через 40 нм встроены молекулы тропонина, а сама цепь охватывает нить тропомиозина. При сокращении мышцы тонкие нити вдвигаются между толстыми нитями. Происходит относительное скольжение нитей без изменения их длины. Этот процесс обусловлен взаимодействием особых выступов миозина - поперечных мостиков с активными центрами, расположенными на актине. Мостики отходят от толстой нити периодично на расстоянии 14,5 нм друг от друга.
В расслабленном состоянии миофибрил молекулы тропомиазина блокируют прикрепление поперечных мостиков к актиновым цепям (Рис.3.а).
 М остик разомкнут. Замыкание. Усиление.
М остик разомкнут. Замыкание. Усиление.


Рис. 3. Процесс активации мостика и генерации усилия в саркомере.
Ионы Са активируют мостики и открывают участки их прикрепления к актину (рис. б).
В результате мостики миозина прикрепляются к актиновым нитям, расщепляются молекулы АТФ и изменяется конфирмация мостиков: их головки поворачиваются внутрь саркомера (Рис.3. в). Это приводит к генерации силы, скольжению актина относительно толстой нити миозина к центру саркомера, что вызывает укорочение мышцы. После окончания активации мостик размыкается и саркомер возвращается в исходное состояние. При укорочении объем саркомера практически не меняется, следовательно, он становится толще, что и подтверждается на снимках поперечного сечения мышц с помощью электронной микроскопии. Каждый цикл замыкание - размыкание сопровождается расщеплением одной молекулы АТФ. Таким образом, актин - миозиновый комплекс
является механохимическим преобразователем энергии АТФ. Рассмотренная структура и последовательность процессов называется моделью скользящих нитей.
Впервые скольжение нитей в саркомере было обнаружено английским ученым Х.Хаксли. Он же сформулировал модель скользящих нитей. Существенный вклад в разработку теории скользящих нитей внес В.И. Дещеревский,
Представленная структурная модель скользящих нитей надежно подтверждена экспериментально и является опытным фактом, на котором основаны все современные теории мышечного сокращения. Эксперементальные данные о микроструктуре мышц были изучены с помощью электронной микроскопии, рентгеноструктурного анализа и метода дифракции синхротронного излучения.
Основные положения модели скользящих нитей:
1. Длины нитей актина и миозина в ходе сокращения не меняются.
2. Изменение длины саркомера при сокращении - результат относительного продольного смещения нитей актина и миозина.
3. Поперечные мостики, отходящие от миозина, могут присоединяться к комплементарным центрам актина.
4. Мостики прикрепляются к актину не одновременно.
5. Замкнувшиеся мостики подвергаются структурному переходу, при котором они развивают усилие, после чего происходит их размыкание.
6. Сокращение и расслабление мышцы состоит в нарастании и последующем уменьшении числа мостиков, совершающих цикл замыкaние - размыкание.
7. Каждый цикл связан с гидролизом одной молекулы АТФ.
8. Акты замыкания-размыкания мостиков происходят не зависимо друг от друга.
На рис. 4. представлена зависимость максимального значения развиваемой силы от степени перекрытия актиновых и миозиновых нитей.

 Возможность саркомера сократиться и развить усилие в большой степени зависит от начальных условий. Если саркомер изначально растянут (его длина 3,65 мкм), то мостики не перекрываются актиновыми нитями и при стимуляции такого элемента усилие не формируется (стрелка 1 на рис. 4, а и фрагмент 1 на рис. 4, б). Если саркомер находится в рабочем начальном состоянии (размер саркомера 2,2 мкм), то при стимуляции он разовьет максимальную силу (стрелка 2, рис. 4, а и фрагмент 2 рис. 4, б). Если начальный размер саркомера слишком короток, генерация усилия уменьшается (стрелка и фрагмент 5).
Возможность саркомера сократиться и развить усилие в большой степени зависит от начальных условий. Если саркомер изначально растянут (его длина 3,65 мкм), то мостики не перекрываются актиновыми нитями и при стимуляции такого элемента усилие не формируется (стрелка 1 на рис. 4, а и фрагмент 1 на рис. 4, б). Если саркомер находится в рабочем начальном состоянии (размер саркомера 2,2 мкм), то при стимуляции он разовьет максимальную силу (стрелка 2, рис. 4, а и фрагмент 2 рис. 4, б). Если начальный размер саркомера слишком короток, генерация усилия уменьшается (стрелка и фрагмент 5).
Рис. 4. Зависимость максимального значения развиваемой силы Р, развиваемой при изометрическом сокращении от начальной длины саркомера l (а) и степени перекрытия актиновых и миозиновых нитей (б).
Мышцы можно представить как сплошную среду, т.е. среду, состоящую из большого числа элементов, взаимодействующих между собой без соударений и находящихся в поле внешних сил. Мышца одновременно обладает свойством упругости и вязкости, то есть является вязко-упругой средой. Для такой среды используются законы классической механики.
Фундаментальными понятиями механики сплошных сред являются деформация, напряжение, упругость, вязкость, а также энергия и температура.
а). Упругость -свойство тел менять размеры и форму под действием сил и самопроизвольно восстанавливать их при прекращении внешних воздействий.
Упругость тел обусловлена силами взаимодействия его атомов и молекул. При снятии внешнего воздействия тело самопроизвольно возвращается в исходное состояние.
б) Вязкость – внутренне трение среды.
в). Вязкоупругость – это свойство материалов твердых тел сочетать упругость и вязкость.
г) Деформация относительное изменение длины: 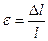 ,
,
где l -начальная длина,  - значение удлинения, может изменять знак.
- значение удлинения, может изменять знак.
д) Напряжение механическое  - мера внутренних сил, возникающих при деформации материала. Для однородного стержня:
- мера внутренних сил, возникающих при деформации материала. Для однородного стержня:  , где S ~ площадь сечения, F - сила,
, где S ~ площадь сечения, F - сила,  приложенная к стержню.
приложенная к стержню.
Упругая деформация возникает и исчезает одновременно с нагрузкой и не сопровождается рассеянием энергии. Для упругой деформации справедлив закон Гука: 
где Е - модуль Юнга, определяемый природой вещества. При растяжении различных материалов, в общем случае,  . При малых растяжениях считают Е=const.
. При малых растяжениях считают Е=const.
Таблица з начений модуля Юнга для различных материалов.
| Материал | Модуль Юнга (Па) |
| Эластин | 
|
| Коллаген | 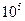
|
| Кость | 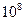
|
| Резина | 
|
| Дуб | 
|
| Сталь | 
|
Эластин - упругий белок позвоночных, находится, в основном, в стенках артерий. Коллаген - волокнистый белок. В мышцах 20 % всех белков – коллаген, он находится также а сухожилиях, хрящах, кости.
В случае вязкой среды напряжение  определяется скоростью деформации d
определяется скоростью деформации d  /dt,
/dt,  , где
, где  - коэффициент вязкости среды.
- коэффициент вязкости среды.
Для вязкоупругой деформации характерна явная зависимость от процесса нагружения во времени, причём при снятии нагрузки деформация с течением некоторого времени самопроизвольно стремится к нулю.
Пассивные механические свойства вязкоупругой среды можно моделировать сочетанием упругих и вязких элементов. Мышцане является ни чисто упругим, ни чисто вязким элементом. Мышца - вязкоупругий элемент.
Пассивное растяжение. На основании расчетных и экспериментальных данных показано, что наиболее простой моделью, дающей достаточно хорошее приближение кмеханическим свойствам мышцы, является трехкомпонентная модель Хилла (Рис. 5).
 Для установления характерной зависимости деформации мышцы во времени при приложении к ней мгновенного воздействия упростим модель (рис. 5). Допустим, что элемент Е2, отсутствует (
Для установления характерной зависимости деформации мышцы во времени при приложении к ней мгновенного воздействия упростим модель (рис. 5). Допустим, что элемент Е2, отсутствует ( ). Тогда для вязкоупругой среды напряжение в системе будет определиться упругой
). Тогда для вязкоупругой среды напряжение в системе будет определиться упругой  с и вязкой составляющими:
с и вязкой составляющими:  + ,
+ , 
Найдем зависимость (t), решая дифференциальное уравнение и считая, что в начальный момент времени t = 0 деформация = 0. B результате получим: 
где величина  называется временем запаздывания.
называется временем запаздывания.
Скорость нарастания деформации максимальна при t = 0: 
Отсюда следует, что чем больше величина вязкой компоненты , тем меньше угол наклона кривой (t).
Деформация (t) растет с убывающей скоростью и асимптотически приближается к стационарному значению  :
: 
Таким образом, уравнение достаточно хорошо описывает процесс нарастания деформации (t), полученный в эксперименте.
В терминах механической модели (рис, 5) и модели скользящих нитей (рис. 3} параллельный упругий элемент Е1, определяет механические свойства внешних мембран клеток (сарколеммы) и внутренних структур - Т-системы и саркоплазматического ретикулума.
Последовательный элемент Е2 определяет упругость актин-миозинового комплекса, обусловленную, прежде всего, местами прикрепления актина к Z-дискам и местами соединения мостиков с активными центрами тонких нитей.
 Вязкий элемент обусловлен скольжением нитей актина относительно миозина. Эта компонента резко возрастает при пассивном режиме мышцы, т. к. в этом случае мостики разомкнуты. Это проявляется в возможности сильного растяжения пассивной мышцы даже при незначительных нагрузках.
Вязкий элемент обусловлен скольжением нитей актина относительно миозина. Эта компонента резко возрастает при пассивном режиме мышцы, т. к. в этом случае мостики разомкнуты. Это проявляется в возможности сильного растяжения пассивной мышцы даже при незначительных нагрузках.
Важно, что уже в этой модели было показано существование вязкой компоненты в мышце, но  физическая природа ее оста вались не ясной.
физическая природа ее оста вались не ясной.
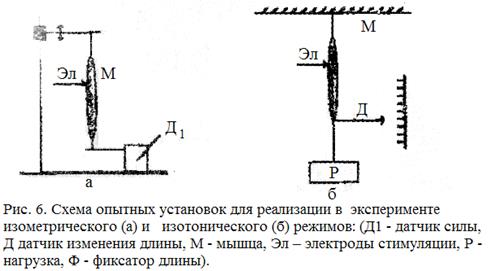
Активное сокращение мышцы. Для исследования характеристик сокращающихся мышц используют два искусственных режима:
1. Изометрический режим, при котором длина мышцы l = const, а регистрируется развиваемая сила F(t).
2. Изотонический режим, при котором мышца поднимает постоянный груз Р = const, а регистрируется изменение ее длины во времени  .
.
При изометрическом режиме с помощью фиксатора (рис 6. а) предварительно устанавливают длину мышцы l. После установки длины на электроды Э подается электрический стимул, и с помощью датчика регистрируется функция F(t). Вид функции F(t) в изометрическом режиме для двух различных длин представлен на рис. 7, а.
Максимальная сила Рп, которую может развивать мышца, зависит от ее начальной длины и области перекрытия актиновых и миозиновых нитей, в которой могут замыкаться мостики: при начальной длине саркомера 2,2 мкм в сокращении участвуют все мостики (см. рис. 4).
Поэтому максимальная сила генерируется тогда, когда мышца предварительно растянута на установке (рис, 6, а) так, чтобы длины ее саркомеров были близки к 2,2 мкм. На рис. 7, а это соответствует начальным длинам двух мышц 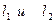 . Но, т. к. количество мостиков в мышце
. Но, т. к. количество мостиков в мышце 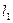 , больше, чем в мышце
, больше, чем в мышце  (
( ), то сила, генерируемая 1г больше.
), то сила, генерируемая 1г больше.
При изотоническом режиме к незакрепленному концу мышцы подвешивают груз Р (рис.6,б). После этого подаётся стимул и регистрируется изменение длины мышцы во времени: . Вид этой функции в изотоническом режиме для двух различных нагрузок показан на рис. 7.б.
Как следует из рис. 7, б, чем больше груз Р, тем меньше укорочение мышцы и короче время удержания груза. При некоторой нагрузке Р = Р0 мышца совсем перестает поднимать груз; это значение. Р0 и будет максимальной силой изометрического сокращения для данной мышцы (рис. 7. а).
Здесь важно отметить, что при увеличении нагрузки угол наклона восходящей части кривой изотонического сокращения уменьшается (рис. 7,б):  <
<  . Другими словами, скорость укорочения с ростом нагрузки падает.
. Другими словами, скорость укорочения с ростом нагрузки падает.
|
|
|
|
|
Дата добавления: 2017-01-14; Просмотров: 1495; Нарушение авторских прав?; Мы поможем в написании вашей работы!