КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Эмбриональная индукция
|
|
|
|
По мере развития организма взаимодействия отдельных клеток сменяются взаимодействиями более крупных элементов зародыша - клеточных комплексов, формирующих структуры, ткани, зачатки органов зародыша. Примером таких влияний служит эмбриональная индукция - взаимодействие элементов развивающегося зародыша, при котором воздействие одного из них направляет (индуцирует) развитие другого. В результате такого взаимодействия запускается цепь морфогенетических (формообразовательных) процессов. Элемент, оказывающий воздействие, назван индуктором. Способность воспринимать индукционное воздействие и отвечать на него определяется как компетенция, а элемент организма, способный реагировать на индукционное воздействие изменением своего развития, назван компетентной тканью. В результате компетентная ткань становится детерминированной (предопределенной) к специфическому типу развития. Далее детерминированное состояние реализуется в процессе дифференцировки (дифференциации).
Следует понимать, что индукционные взаимодействия осуществляются на основе межклеточных, без которых не обходится ни один этап развития.
Феномен эмбриональной индукции был открыт немецким эмбриологом Г. Шпеманом и его ученицей Г. Мангольд в 1921 г. в серии экспериментов по изучению свойств материала хордомезодермы. Для того чтобы иметь возможность проследить за судьбой клеток при трансплантации, были использованы два вида тритонов, отличающихся по окраске эмбриональных тканей: гребенчатый тритон, клетки которого не содержат пигмента, и обычный тритон с пигментированными клетками. Участок дорзальной губы бластопора, содержащий материал хордоме-зодермы, зародыша гребенчатого тритона на стадии ранней гаструлы пересаживали в (под) боковую или брюшную эктодерму обыкновенного приблизительно той же стадии развития. У зародыша-реципиента в месте пересадки наблюдалось образование второго комплекса осевых органов (хорды, нервной трубки и сомитов). В некоторой доле случаев развитие завершалось формированием дополнительного зародыша (рис. 8.31). По распределению неокрашенных и пигментированных клеток было установлено, что почти вся нервная трубка и значительная часть мезодермы возникли из тканей реципиента, а пересаженная хор-домезодерма образовала, как и следовало ожидать, хорду, часть мезодермы, а также небольшой участок нервной трубки.
Описанное явление получило название первичной эмбриональной индукции. Зачаток хордомезодермы, локализованный в дорзальной губе бластопора, был назван первичным эмбриональным индуктором. Эктодерма, воспринимающая воздействие и отвечающая формированием нервной трубки в этом эксперименте представляет собой компетентную ткань.
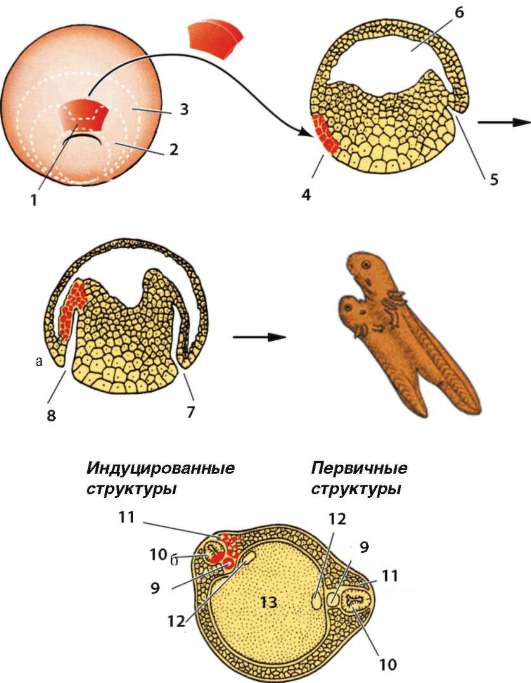
Рис. 8.31. Эксперимент Г. Шпемана по пересадке спинной губы бластопора от зародыша-донора зародышу-реципиенту: а - схема опыта; б - поперечный срез на стадии закладки двух комплексов осевых органов. 1 - спинная губа бластопора; 2 - презумптивная мезодерма; 3 - презумптивная хорда; 4 - материал донора; 5 - инвагинация; 6 - бластоцель; 7 - первичная инвагинация; 8 - вторичная инвагинация; 9 - хорда; 10 - нервная трубка; 11 - мезодерма; 12 - полость кишки; 13 – энтодерма
В ходе дальнейших исследований было показано, что ткань индуктора не сразу приобретает способность направлять формирование всего спектра структур, который образуется под ее влиянием. Наблюдается созревание способности к индукции - постепенное приобретение каждой частью индуцирующей ткани возможности оказывать воздействие на формирование определенных элементов зародыша. Если пересадить дорзальную губу ранней гаструлы, то индуцируется развитие структур переднего мозга, если же пересадить дорзальную губу поздней гаструлы, то развиваются спинной мозг и мезодермаль-ные ткани (рис. 8.32).
Осуществление индукции возможно лишь при условии, что клетки реагирующей ткани способны воспринять воздействие, т.е. являются компетентными. Только в этом случае они отвечают образованием соответствующих структур. Компетенция ткани также возникает на определенной стадии развития. Клетки реагирующей ткани должны пройти определенные фазы развития, прежде чем они приобретут способность к дифференцировке под влиянием сигналов индуктора. Состояние компетенции к воздействию определенного индуктора сохраняется ограниченное время. Затем может появиться компетенция к другому индуктору. Для каждого индуктора также характерно наличие определенного периода функциональной активности.
В эксперименте на амфибиях показано, что компетенция к образованию нервной ткани у амфибий возникает с начала гаструляции и затрагивает всю эмбриональную эктодерму. К концу этой стадии компетенция прекращается. Время контакта между хордой и нейроэктодер-мой при первичной индукции должно быть не менее 4 ч. При меньшем по времени индуцирующем воздействием формирования нейральных структур не происходит.

Рис. 8.32. Результаты пересадки дорзальной губы бластопора на стадиях ранней (а) и поздней (б) гаструлы (объяснение в тексте)
Пересадка материала дорзальной губы бластопора на стадии нейру-ляции не приводит к формированию дополнительной нервной трубки. Это объясняется тем, что эктодерма на указанной фазе развития уже не способна отвечать на сигналы данного индуктора. Однако она становится компетентна в отношении иных индукторов. Например, на индуцирующее действие глазного пузыря отвечает образованием хрусталика. Задний мозг сходным образом может индуцировать образование из прилегающей к нему эктодермы слухового пузырька.
Кроме вышеперечисленного для эффективного ответа на индуцирующее влияние необходимо наличие в компетентной ткани определенного, минимального числа клеток, т.е. требуется некоторый «порог массы». Одиночные клетки не воспринимают действие индуктора. Если же их число превышает «порог массы» и клетки обладают минимальной организацией, то количество образуемых структур из возможного спектра для данной конкретной индукции зависит от объема реагирующей ткани. Чем больше в ней клеток, тем активнее ее реакция. При этом для оказания индуцирующего воздействия достаточно лишь одной клетки индуктора.
Во всех классах хордовых индукционные взаимодействия между хор-домезодермальным и нейральным зачатками подобны таковым у амфибий. У зародышей амниот (птиц, рептилий и млекопитающих) зачаток хордомезодермы локализован в области гензеновского узелка. Поэтому второй зародыш сходным образом «организуется» благодаря воздействию спинной губы бластопора, а у птиц и млекопитающих подобное действие оказывает гензеновский узелок. Интересно, что у ланцетника и круглоротых активен только туловищный индуктор, стимулирующий формирование нервной трубки (спинного мозга), а головной индуктор не действует. Это, безусловно, связано с отсутствием головного мозга у бесчерепных и слабым его развитием у круглоротых. У костистых рыб уже присутствуют оба индуктора.
В целом формирование гомологичных структур в группах эволю-ционно родственных организмов происходит под контролем сходных индукций. Так, для формирования придатков кожи необходимо стимулирующее влияние мезодермы на эпидермис кожи. Показано, что начальные этапы формирования кожных придатков у амниот можно индуцировать дермой зародышей других классов, например дерма ящерицы стимулирует развитие волос в эпидермисе мыши. Это свидетельствует о том, что эмбриональная индукция как один из важнейших механизмов развития имеет большое эволюционное значение, а сами индукторы зачастую эволюционно консервативны.
После открытия явления первичной эмбриональной индукции были предприняты неоднократные попытки идентифицировать индуцирующие молекулы, выделяемые первичным организатором, определить их свойства и механизм действия. В 1932 г. группа исследователей, возглавляемая Г. Шпеманом, экспериментально продемонстрировала химическую природу индуцирующего сигнала, вызывающего формирование нейральных структур. Вскоре выяснилось, что индукцию могут вызывать разнообразные убитые ткани, вытяжки из различных живых тканей беспозвоночных и позвоночных животных, а также растений, несколько классов химических соединений (белки, нуклеопротеины, стероиды и даже неорганические вещества).
Новый этап исследований молекулярных механизмов эмбриональной индукции начался примерно 20 лет назад, когда благодаря прогрессу молекулярной биологии оказалось возможным связать индукционные процессы, как и вообще клеточную дифференцировку, с активацией или репрессией работы определенных генов, ответственных за синтез специфических белков.
Было установлено, что на ранних стадиях эмбриогенеза в зародыше синтезируются белки семейства ВМР (от англ. bone morphogenetics proteins - морфогенетические белки, получаемые из костного мозга), входящие в надсемейство TGF-β - трансформирующих факторов роста β. Их концентрация наивысшая на вентральной стороне зародыша. Белки секретируются в межклеточное пространство, связываются с мембранными рецепторами эмбриональных клеток и препятствуют их диффе-ренцировке в нервную ткань и другие производные осевых зачатков, позволяя развитие только в сторону покровной эктодермы. Для осуществления формирования нервной трубки (нейральной дифференци-ровки) взаимодействие ВМР с рецепторами мембран клеток-мишеней должно быть предотвращено.
Клетки шпемановского организатора - хордомезодермы - секре-тируют в межклеточное пространство белки chordin и noggin. Их функция состоит в том, чтобы связывать молекулы ВМР в межклеточном пространстве, препятствуя их взаимодействию с мембранными рецепторами клеток. В отсутствие ВМР клетки дорзальной эктодермы дифференцируются в нервную ткань (рис. 8.33). Таким образом, реализуется «индукция по умолчанию», поскольку данная дифференцировка не требует дополнительных стимулирующих воздействий, а нуждается лишь в блокировании ВМР, и именно его осуществляет шпемановский индуктор.
Это открытие привело к существенному пересмотру традиционных представлений о первичной индукции. Ранее считалось, что базисный путь дифференцировки эмбриональных клеток, не требующий индукционных влияний, - их развитие в покровную эктодерму.
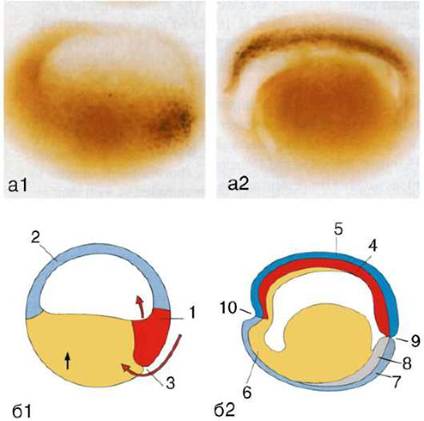
Рис. 8.33. Локализация мРНК белка noggin в ткани зародыша амфибии, выявленная методом гибридизации in situ (черные точки): а - фотографии; б - соответствующие схемы. а1, б1 - бластула; а2, б2 - гаструла. 1 - шпемановский организатор; 2 - презумптивная эктодерма; 3 - место начала инвагинации (дорзальная губа бластопора); 4 - зачаток хорды (хордомезодерма); 5 - ней-роэктодерма; 6 - энтодерма; 7 - эктодерма; 8 - мезодерма; 9 - бластопор; 10 - место образования будущего рта
Подразделение нервной системы на отделы также осуществляется путем «индукции по умолчанию». Выяснено, что в межклеточном пространстве на стадии гаструлы присутствуют белки семейства Wnt. Оказалось, что если не препятствовать их связыванию с рецепторами клеток презумптивной (предполагаемой) нейральной эктодермы, то вся нервная пластинка развивается в спинной мозг. Вещества семейства Wnt связываются в межклеточном пространстве белками ucrberus и Dickkopf, которые секретируются передней частью хордомезодермы - прехор-дальной пластинкой. Следствием такого взаимодействия становится активация в клетках, соответственно, передней части нервной пластинки определенных генов, среди которых ОТХ-2, anf и другие, что и приводит в результате к формированию головного мозга и его отделов.
Однако механизмы индукции не определяются только лишь включением и выключением определенных генов. Как и любой процесс в организме, регуляция индуктивных взаимодействий осуществляется на нескольких уровнях, она многогранна и к настоящему времени еще далеко не полностью изучена. Так, несмотря на многочисленные эксперименты, не удалось с достоверностью обнаружить химический фактор, выделяемый глазным бокалом и необходимый для индукции хрусталика, хотя его существование утверждается рядом исследователей.
Как было показано в предыдущем разделе, межклеточные взаимодействия, лежащие в основе индукции, могут осуществляться не только вследствие выделения клеткой каких-либо факторов, но и при непосредственном межклеточном контакте, а также через матрикс. Все это справедливо и для индуктивных взаимодействий. Например, установлено, что для развития метанефрогенной мезенхимы формирующейся почки под действием чужеродного индуктора (нервной ткани) необходим непосредственный контакт между клеточными отростками индуктора и реагирующей ткани. Роль компонентов внеклеточного матрикса в индуктивных процессах показана в том числе и в экспериментах со стволовыми клетками. Одна и та же стволовая клетка при добавлении в среду коллагена IV типа может дать начало эпителиальным клеткам, при добавлении фибронектина и коллагена I - соединительной ткани, а коллагена II - хрящу.
В 50-60-е гг. ХХ в. голландский эмбриолог П. Ньюкоп продемонстрировал, что первым индуцирующим событием в развитии зародыша является не воздействие хордомезодермы на дорзальную эктодерму на стадии ранней гаструлы, а осуществляемая на стадии бластулы индукция энтодермой (клетками, расположенными на вегетативном полюсе зародыша) преобразования смежных клеток в хордомезодермальную закладку (рис. 8.34). По сути дела, данное событие и есть истинно первичная эмбриональная индукция, что было подтверждено экспериментально. После удаления у зародыша-бластулы вегетативных клеток образования хорды и ряда мезодермальных структур не происходило. Опыты по рекомбинации клеток зародышей показали, что наиболее дорзальные бластомеры вегетативного полюса индуцируют развитие хорды и сомитов, а прочие клетки этого полюса определяют образование вентральных мезодермальных структур, прежде всего боковой пластинки. Таким образом, шпемановская индукция базируется на прошедшей перед этим индукции мезодермы. Но так как открытие Г. Шпеманом было сделано раньше, за ним закрепилось наименование первичная эмбриональная индукция. Все индукции, осуществляемые после нее, называют вторичными, третичными и т.д.
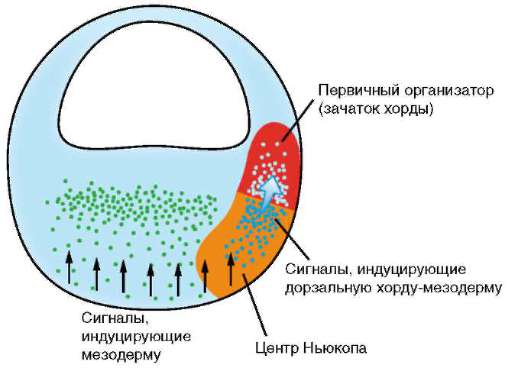
Рис. 8.34. Схематическое изображение влияний, реализуемых при индукции Ньюкопа на стадии бластулы
Описанные эксперименты могут быть расценены не как проявление индукционных влияний, а как один из примеров межклеточных взаимодействий. Однако они, несомненно, доказывают преемственность этих механизмов развития.
Действительно, начиная со стадии бластулы, уже на самых ранних этапах гаструляции выявляется выраженная кооперативность клеточного поведения, когда действуют не отдельные клетки, а клеточные группы, составляющие зачатки структур, тканей и органов особи. Гетерогенность клеточных популяций, взаимодействие между собой отличающихся друг от друга комплексов клеток - основа, на которой возникает дифференциальная активность генов на тканевом уровне, что приводит к дифференциации структур и формированию органов. Под дифференциацией понимают возникновение в процессе развития в отдельном участке (структуре, части) организма морфологических и функциональных различий. Дифференциация структур базируется на дифференцировке клеток развивающегося организма.
В настоящее время установлено, что процессы эмбриональной индукции представляют собой каскад взаимодействий, которые определяют последовательное формирование структур и органов зародыша, его полноценное развитие. По современным данным, материальная основа этого каскада закладывается еще в оогенезе. Так, у амфибий непременной составной частью инициации данного процесса является описанный в п. 8.2.6 поворот оплодотворения.
В ходе роста ооцита амфибий поблизости от его вегетативного полюса синтезируется большое число белков, впоследствии участвующих в индукционных процессах, в том числе факторы семейства Wnt-1 и надсемейства TGF-β. мРНК для всех этих белков были синтезированы в оогенезе на хромосомах типа ламповых щеток. Ньюкоповская индукция опосредуется, в частности, белком Vg1, принадлежащим к над-семейству TGF-β. Другие участники этого процесса - белок dishevelled, синтезируемый в вентральной области яйцеклетки в период оогенеза, и β-катенин, который исходно распределен в цитоплазме зиготы более или менее равномерно. Вскоре после оплодотворения β-катенин подвергается ферментативному расщеплению, однако на дорсальной стороне зародыша активность расщепляющего фермента подавляется белком dishevelled (Dsh), который перемещается в эту область зародыша именно в результате поворота оплодотворения. Вследствие этого на дорсальной стороне β-катенин сохраняется и по мере делений дробления перемещается в клеточные ядра бластомеров (рис. 8.35). Роль β-катенина состоит в том, что он связывается с промоторами определенных генов, активируя их. Кодируемые ими белки, в свою очередь, оказывают аналогичное влияние на другие гены, и в результате запускается целая цепь генов, активирующих друг друга и участвующих в реализации индукционного каскада. Продукт одного из активированных таким образом генов - goosecoid - воздействует в клетках шпемановского организатора на гены, кодирующие уже знакомые нам белки chordin и noggin, т.е. факторы первичной эмбриональной индукции.
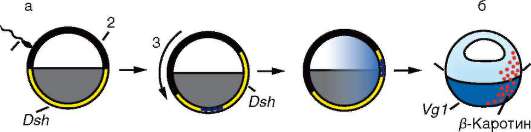
Рис. 8.35. Схема взаимодействия генов, инициированного поворотом оплодотворения у амфибий: а - последовательные стадии изменения распределения белка dishevelled (Dsh) в результате оплодотворения, б - распределение в цитоплазме зиготы; 1 - сперматозоид, 2 - кортикальный слой цитоплазмы, 3 - поворот оплодотворения, 4 - брюшная сторона зародыша, 5 - спинная сторона
Как было сказано выше, дифференциация большинства структур и органов в процессе развития зависит от предшествующих индукционных событий. При этом в ходе их реализации наблюдается последовательная смена индукторов и состояний компетентности. Именно таким образом взаимодействие между зачатком хордомезодермы и дорзаль-ной эктодермой инициирует цепь формообразующих процессов. Сформированные в результате первичной эмбриональной индукции хорда и нервная трубка необходимы при дифференцировке клеток мезодермы для образования сомитов и для дифференцировки хрящевых клеток из дерматомов. Наличие сомитов в свою очередь обязательно для нормального формирования отделов кишечной трубки. Присутствие дорзальной энтодермы кишки оказывает индуцирующее влияние на развитие кроветворных участков мезодермы и т.д.
Индукция носит не только каскадный, но и переплетающийся, взаимный (реципрокный) характер, что может быть проиллюстрировано на примере формирования глаза (рис. 8.36.). Вырост переднего мозга - глазной пузырь - инициирует образование хрусталиковой плакоды из лежащей над ним эктодермы. Далее направление индукции меняется и, сформировавшись, хрусталиковая плакода, в свою очередь, вызывает изменения в глазном пузыре, передняя стенка которого инвагинирует, и пузырь превращается в двустенную чашу - глазной бокал. Одновременно с этим два слоя глазного бокала начинают дифференцироваться в разных направлениях: внутренний становится сетчаткой, а наружный - пигментным эпителием. Под действием сетчатки, которая на этом этапе становится индуктором, из хрусталиковой плакоды образуется хрусталик. Последний вызывает формирование роговицы из прилежащей к нему эктодермы и оказывает направленное действие на окончательную дифференцировку клеток глазного бокала. Роговица в свою очередь также приобретает свойства индуктора и участвует в формировании век. Интересно отметить, что образующийся хрусталик выделяет вещества, препятствующие развитию еще одного хрусталика.
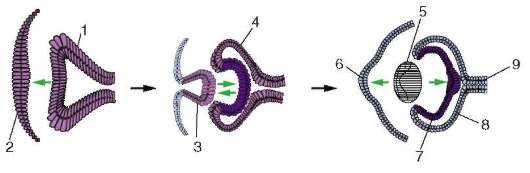
Рис. 8.36. Схема, иллюстрирующая реципрокный характер индукции: 1 - глазной пузырь, 2 - покровная эктодерма, 3 - формирующийся хрусталик, 4 - глазной бокал, 5 - сформированный хрусталик, 6 - роговица, 7 - нейральный слой сетчатки, 8 - пигментный слой сетчатки, 9 - зрительный нерв
Интересен факт, что в индукции может наблюдаться «кумулятивный» эффект, т.е. в индукции той или иной структуры может участвовать не одна, а несколько тканей. Например, глазной бокал служит главным, но не единственным индуктором хрусталика. В ходе развития презум-птивный хрусталик, т.е. эпидермис, из которого затем должен развиться хрусталик, во время гаструляции лежит над энтодермой будущей глотки, первого индуктора хрусталика. Затем под этим эпидермисом оказывается сердечная мезодерма, которая также действует как индуктор. И только позднее, во время нейруляции на переднем конце нервной трубки выпячиваются глазные пузыри, образующие глазной бокал и сетчатку, являющуюся главным индуктором хрусталика (рис. 8.37). Путем удаления той или иной из индуцирующих тканей определили степень участия каждой из них в индукции хрусталика. Оказалось, что при удалении сетчатки глазного бокала у 42% зародышей амфибий все же формировались хрусталики и, следовательно, энтодерма и мезодерма в сумме обладают почти таким же индуцирующим действием, как и сетчатка глазного бокала. Полагают, что многочисленность индуцирующих тканей может иметь решающее значение для точного установления места формирования органа. Кроме того, последовательные индукции могут играть важную стабилизирующую роль в развитии, обеспечивая нормальное течение органогенеза, даже если один из компонентов индуцирующей системы не сумеет произвести сигнал нужной силы.
Различают гетерономную и гомономную индукцию. К гетерономной относят случаи, при которых одна структура зародыша индуцирует формирование иной структуры (хордомезодерма индуцирует появление нервной трубки и всего зародыша в целом). Гомономная индукция заключается в том, что индуктор побуждает окружающий материал к развитию в том же направлении, что и он сам. Например,область нефротома, пересаженная другому зародышу, способствует развитию окружающего материала в сторону формирования головной почки, а прибавление в культуру фибробластов сердца маленького кусочка хряща влечет за собой процесс образования хряща.
Рис. 8.37. Последовательные индукционные взаимодействия, необходимые для образования хрусталика у зародыша амфибии: I - ранний зародыш; II - гастру-ла; III - нейрула; IV - стадия хвостовой почки; V - личинка; VI - взрослая особь. 1 - хрусталиковая плакода; 2 - хрусталиковый пузырек; 3 - хрусталико-вые волокна. а - энтодерма; б - сердечная мезодерма; в – сетчатка
Установлено, что индуктивные взаимодействия наиболее характерны для онтогенеза животных, характеризующихся регуляционным типом развития. Что же касается организмов с явно выраженным мозаичным онтогенезом, то у них явления типа эмбриональной индукции имеют меньшее значение, однако также оказывают определенное воздействие на клеточную дифференцировку. Например, ряд аналогичных процессов наблюдается в формировании нервной системы.
Так, у асцидий, на стадии 8 бластомеров, когда уже все основные зачатки предопределены, проводили некоторые перемещения клеток. Материал хордомезодермы и основная часть нейрального материала у них локализованы в заднем вегетативном бластомере. Небольшая часть нейрального материала, формирующего головной ганглий, находится в заднем анимальном бластомере, расположенном над задним вегетативным (рис. 8.38). Для проверки индукционных взаимодействий между ними анимальный ярус бластомеров поворачивали на 180° так, чтобы задний анимальный бластомер терял контакт с задним вегетативным. Головной ганглий не развился нигде. Это означает, что для развития головного ганглия необходимо индукционное влияние на задний ани-мальный бластомер со стороны заднего вегетативного. Кроме того, очевидно, что задний анимальный бластомер не обладает автономностью развития, но только он компетентен к восприятию воздействия со стороны заднего вегетативного бластомера, содержащего хордомезодер-мальный зачаток.
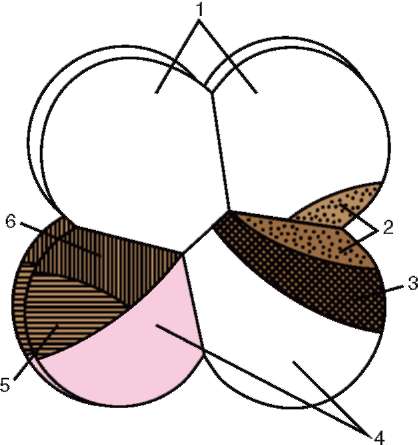
Рис. 8.38. Карта презумптивных зачатков у зародыша асцидий на стадии восьми бластомеров: 1 - эпидермис; 2 - нервная пластинка; 3 - хорда; 4 - энтодерма; 5 - сомиты, 6 – мезодерма
Полноценная эмбриональная индукция зависит от того, насколько точно соответствует в развитии время созревания индуктора и компетентной ткани. В нормальных условиях компетентная ткань способна отвечать формообразованием в момент стимулирующего импульса от индуктора. Рассогласования во времени созревания индуктора и компетентной ткани нарушают ход соответствующих морфогенетических процессов. Мутации, вызывающие такие рассогласования, вероятно, распространены довольно широко.
Так, становление пигментации у амфибий определяется взаимодействием эпидермиса (индуктора) и ткани нервного гребня, который служит источником меланобластов, мигрирующих субэпидермально под влиянием индуктора. Одна из мутаций в гомозиготном состоянии резко ослабляет окраску аксолотля, так что лишь спина животного слегка окрашена (так называемая белая раса аксолотлей). Экспериментально показано, что отсутствие окраски определяется рассогласованием во времени созревания двух взаимодействующих закладок, составляющих единую индукционную систему. При трансплантации кусочков презум-птивного эпидермиса между зародышами аксолотлей белой расы разного возраста обнаружено, что при некоторых сочетаниях возраста донора и реципиента в трансплантате развивается пигментация.
Исследования, выполненные к настоящему времени, позволили сделать вывод, что большинство индукционных процессов, особенно на более поздних стадиях органогенеза, являются пермиссивными. Это означает, что индуктор лишь запускает процесс дифференцировки, а его результат определяется свойствами компетентной ткани. В этом случае индуцирующий стимул как бы высвобождает ответ, уже предопределенный в клетках реагирующей ткани. Так, например, формирование конечности может быть индуцировано пересадкой слухового пузырька, носовой плакоды или гипофиза.
Изучение эмбриональной индукции, ее молекулярных и клеточных механизмов активно продолжается. Кроме того, это явление представляет большой теоретический интерес, т.к. позволяет оценить взаимоотношение таких процессов, как детерминация, дифференцировка и морфогенез.
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 11142; Нарушение авторских прав?; Мы поможем в написании вашей работы!