КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Локальные механизмы дифференцировки и детерминации
|
|
|
|
Под экспрессией гена понимают синтез в клетке функционально активной формы белка, кодируемого данным геном. Процесс реализации генетической информации у эукариот к настоящему времени достаточно хорошо изучен, однако регуляция этого многоступенчатого процесса очень сложна и неоднозначна (см. главу 2). Данные, накопленные в результате многолетней и кропотливой работы многих исследователей, позволяют выделить следующие возможные уровни регуляции биосинтеза белков, а, следовательно, и дифференцировки клеток, основанной, прежде всего, на приобретении клетками биохимических различий: регуляция путем соматических мутаций, регуляция транскрипции, регуляция процессинга мРНК и транспорта мРНК в цитоплазму, регуляция трансляции, регуляция на посттрансляционном уровне.
К регуляции путем соматических мутаций могут быть отнесены случаи качественного и количественного изменения генетического материала, происходящие в ходе развития в отдельных соматических клетках. Помимо элиминации хромосом, амплификации генов, образования политенных хромосом (см. п. 8.2.5.1), примером может служить перестройка генов иммуноглобулинов, в результате которой в организме человека иммунокомпетентные клетки могут синтезировать широкий спектр различных белков-антител. В организмах высших животных, в том числе человека, существует более миллиона клонов В-лимфоцитов, различающихся по продуцируемым антителам (иммуноглобулинам). Иммуноглобулины состоят из так называемых легких и тяжелых аминокислотных цепей. Гены для легких цепей содержат 2 вариабельных сегмента ДНК (V и J) и константный сегмент С. Сегмент V включает около 250 различных нуклеотидных последовательностей, а сегмент J - 4 таких последовательности. На нитях ДНК еще недифференцированных клеток участки V, J и С пространственно разделены. В эмбриональном развитии в ходе дифференцировки Б-лимфоцитов промежуточная ДНК элиминируется, и любая из V-последовательностей может сблизиться с любой из J-последовательностей, а их комбинация - с константным С-сегментом, т.е. происходят перемещения (транслокации) нуклеотидных последовательностей ДНК. Таким образом, может быть образовано около 1500 различных комбинаций генов. Гены для тяжелых цепей содержат вариабельные сегменты V, D и J, состоящие, соответственно, из 500, 15 и 4 последовательностей, а также константный участок С (рис. 8.24). Их комбинирование дает около 30 000 вариантов. При синтезе конкретного иммуноглобулина объединяются белки, кодируемые одним из генов легких цепей и одним из генов тяжелых цепей, что определяет возможность продуцировать около 100 млн различных типов антител.
|
|
|
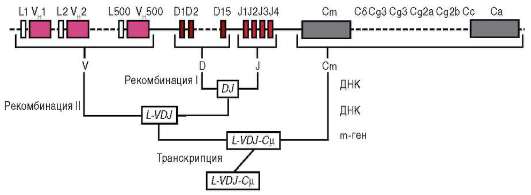
Рис. 8.24. Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и С)
Подобные механизмы, в отличие от обсуждаемых далее, являются скорее исключением и обнаруживаются либо у отдельных видов животных, либо на определенных стадиях развития, либо обеспечивают реализацию конкретной внутриорганизменной задачи.
Как было установлено, в подавляющем большинстве случаев все соматические клетки организма имеют идентичный по количеству и содержанию набор ДНК. Наблюдаемая при их дифференцировке избирательная активность генов регулируется на разных стадиях процесса реализации генетической информации - от транскрипции до посттрансляционных изменений образованных полипептидных цепей - и базируется на действии многообразных механизмов. Экспрессия одного и того же гена может подвергаться действию различных регулирующих механизмов.
Регуляция транскрипции обеспечивает синтез первичных транс-криптов (пре-мРНК) только на определенных структурных генах. Некоторые примеры избирательной транскрипции обсуждались ранее - это синтез мРНК на выпетлившихся участках хромосом ооцита типа «ламповых щеток», а также на вздутых участках - пуфах - политен-ных хромосом в ядрах клеток слюнных желез некоторых насекомых (см. п. 2.4.3.4).
|
|
|
Наборы транскрибируемых генов отличаются в разных клетках на разных этапах развития, что и определяет направление их дифференцировки. В многомодульной регуляции транскрипции в эукариотических клетках принимает участие ряд нуклеотидных последовательностей ДНК, которые выполняют сервисные и регуляторные функции (см. п. 2.4.5.5). Прежде всего, это расположенные в непосредственной близости от кодирующих последовательностей гена участки ДНК: промоторы и операторы. Промоторы связывают РНК-полимеразу, комплекс общих транскрипционных факторов и специфические факторы транскрипции, операторы взаимодействуют с белками-регуляторами и веществами небелковой природы - эффекторами. Наличие нескольких промоторов в одном гене обусловливает альтернативную транскрипцию, т.е. образование различных форм мРНК при инициации считывания с разных промоторов. Так, в гене, кодирующем белок дис-трофин, имеется 8 промоторов, с которых происходит альтернативная транскрипция в разных тканях (сердечных и скелетных мышцах, эмбриональных нейронах, коре головного мозга, сетчатке глаз), что приводит к образованию в этих тканях различных изоформ белка. Обсуждается возможность формирования разных мРНК, транскрибируемых с одного гена, за счет использования альтернативных терминаторов (рис. 8.25).
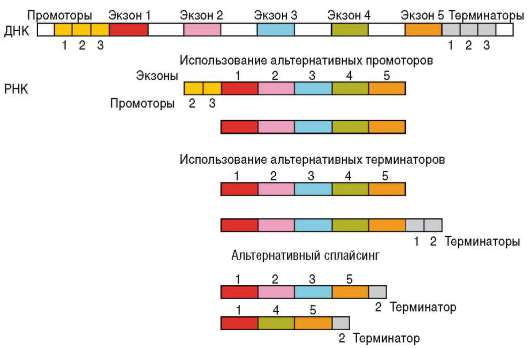
Рис. 8.25. Схема возможных механизмов синтеза различных мРНК с одного гена
Помимо этого имеются области ДНК, называемые энхансерами. Они могут располагаться на значительном расстоянии от регулируемых генов (за тысячи нуклеотидов) и контролировать работу не одного конкретного, а целой группы доступных для них генов. Энхансеры связываются с комплексами белков, которые в зависимости от своего состава могут либо усиливать, либо подавлять транскрипцию данного структурного гена. Воздействие энхансера на конкретный ген осуществляется благодаря изгибу расположенного между ними участка ДНК, в результате чего комплекс энхансер-белки устанавливает непосредственный контакт со структурным геном (см. рис. 2.36). Изгиб становится возможен вследствие деконденсации участка ДНК, расположенного между энхансером и контролируемым им геном.
|
|
|
К процессам, регулирующим активность генов на уровне транскрипции, следует отнести также метилирование-деметилирование различных участков ДНК. Метилирование - присоединение метильной группы к цитозину, наблюдаемое в том случае, если рядом с ним находится гуанин, т.е. в составе динуклеотидов ЦГ (CpG). У млекопитающих в соматических клетках взрослого организма метилирование ДНК обычно происходит в последовательностях длиной 1000-2000 пар - CpG-островках, в которых содержание динуклеотидов ГЦ в 10-20 раз выше, чем в среднем по геному. CpG-островки присутствуют в 5' регуля-торных областях многих генов. Их метилирование препятствует взаимодействию регуляторных белков (факторов транскрипции) с промотором и блокирует активность генов. Обратный процесс - деметилирование приводит, соответственно, к деблокированию активности генов.
Одно из наиболее значимых в последние годы открытий в области регуляции генной экспрессии в раннем периоде онтогенеза млекопитающих - установление феномена перепрограммирования генома. На стадии дробления от зиготы до бластоцисты отмечается практически тотальное деметилирование генома, (за исключением импринтированных локусов (см. п. 4.1.1)), в результате чего происходит активация генов. Предполагается, что подобные преобразования генома существенны для обеспечения тотипотентности зиготы.
В период имплантации, когда наблюдается обособление зародышевых и внезародышевых листков (групп клеток), запускается процесс установления тканеспецифичного метилирования. При этом уровень метилирования ДНК в клетках трофобласта возрастает незначительно, в то время как ДНК клеток-производных внутренней клеточной массы, дающих начало эмбриональным структурам, подвергается существенному метилированию. Возрастающая плотность метилирования коррелирует с последующим сужением спектра возможных путей дифференцировки клеток. Показано, что рисунок метилирования оказывается специфическим для данного типа клеток и способствует поддержанию устойчивости его дифференцировки. У человека за процесс метилирования ДНК отвечают три фермента, называемые ДНК-метилтрансферазами (DNMT1, DNMT3a, DNMT3b). Предполагается, что DNMT3a и DNMT3b - это метилтрансферазы, которые осуществляют формирование рисунка метилирования ДНК на ранних стадиях развития. DNMT1 является, предположительно, ферментом, поддерживающим метилирование ДНК на более поздних стадиях развития организма и отвечающим за присоединение метильной группы на комплементарной цепи при репликации ДНК дочерней клетки.
|
|
|
Установлено, что изменение доступности промоторов для белков, участвующих в транскрипции, и, следовательно, обеспечение избирательной экспрессии генов достигается также благодаря ацетилирова-нию гистонов. Гистоны целенаправленно модифицируются с помощью ферментов ацетилтрансфераз на тех промоторах, которые требуется активировать. Деацетилирование гистонов, в частности Н4, ремоделирует структуру хроматина, повышая степень его компактизации, что приводит к репрессии транскрипции.
Еще один механизм избирательной транскрипции структурных генов в дифференцирующихся клетках связан с пространственной организацией хромосом в интерфазных ядрах. Так, согласно некоторым данным, при дифференцировке В-лимфоцитов гены CD2, CD4, CD8-alpha, CD19, CD45-lambda5 включаются в состав гетерохроматина, в результате чего их экспрессия подавляется. Связь неактивных генов с гетерохромати-ном осуществляется с помощью белка Ikaros, который специфически связывается с промоторами генов и тем самым «рекрутирует» их в состав гетерохроматина.
Другой недавно обнаруженный фактор, влияющий на активность и, возможно, на специфичность транскрипции - размер петлеобразных участков ДНК - доменов, возникающих при прикреплении молекулы к ядерному матриксу (см. п. 2.4.3.2). У шпорцевой лягушки до 12-го деления дробления зародыша транскрипции на его генах не происходит, и петли хроматина не имеют постоянных точек фиксации на ядерном матриксе. С началом экспрессии собственного генома места их прикрепления точно фиксируются. По ходу развития размер хроматиновых петель, как правило, увеличивается. Кроме того, во всех клетках выявлены одинаковые петли ДНК, что позволяет сделать вывод о сохранении в ряду клеточных делений специфической организации доменов, разделенных зонами прикрепления. Предполагается, что при образовании петель могут фиксироваться позиции различных регуляторных элементов и их мишеней - структурных генов, способствуя их взаимодействию либо, наоборот, исключая его.
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 1007; Нарушение авторских прав?; Мы поможем в написании вашей работы!