КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Гетерогенность яйцеклетки как основа дифференцировки
|
|
|
|
Регуляция процессинга РНКранее обозначалась как посттранскрипционная. Считалось, что она осуществляется лишь после окончания транскрипции. По современным данным, однако, процессы «созревания» пре-мРНК протекают еще во время самой транскрипции - котранскрипционно.
Один из широко используемых механизмов в процессинге РНК - альтернативный сплайсинг. Как известно, только что транскрибированная молекула пре-мРНК состоит не только из участков, несущих генетическую информацию (экзонов), но и из некодирующих «вставок» (интронов). Еще в ходе транскрипции интроны удаляются из новосинтезированной мРНК. Оставшиеся экзоны могут соединяться в различных комбинациях, в результате чего из одной молекулы пре-мРНК может образоваться несколько типов более коротких молекул мРНК, кодирующих различные белки (изоформы белка) (см. п. 2.4.5.5, см. рис. 8.25). Было показано, например, что в результате альтернативного сплайсинга первичного транскрипта гена каль-цитонина могут быть образованы различные мРНК. В результате в клетках щитовидной железы он экспрессируется в виде гормона каль-цитонина, а в клетках гипофиза - в виде нейропептида CGRP, и эти пептиды будут участвовать в дифференцировке совершенно различных типов клеток.
Анализ сотен миллионов фрагментов РНК из разных тканей и органов человека показал, что до 60%, а по некоторым данным, до 94% генов подвергаются альтернативному сплайсингу, причем в разных тканях производятся разные наборы изоформ белков. Благодаря альтернативному сплайсингу разнообразие белков в организме млекопитающих значительно выше, чем у низших животных, хотя количество генов у тех и других примерно одинаково.
Интересно, что среди генов, сплайсинг которых отличается наиболее строгой тканеспецифичностью (в одних тканях всегда или почти всегда синтезируется только одна изоформа, в других - другая) повышена доля генов-регуляторов индивидуального развития, обмена веществ, межклеточных взаимодействий и передачи сигналов. Это именно те функции, от которых зависят структурные и функциональные различия между клетками и тканями.
В ходе формирования функционально активной мРНК может осуществляется также редактирование (эдитинг) РНК. Суть его состоит во внесении изменений в молекулу РНК - замена оснований, вырезание и вставка нуклеотидов, которые позволяют синтезировать иные полипептиды на отредактированных матрицах. Хорошо изучен процесс редактирования мРНК аполипопротеина В (АроВ) млекопитающих, участвующего в транспорте холестерина и триглицеридов. Обнаружены две формы AроB, кодируемые одним и тем же геном и образующиеся в результате редактирования AроB-мРНК. Ген AроB человека содержит 29 экзонов, и его общая длина составляет 43 тыс. п.о. Длина мРНК этого гена, кодирующей ApoB100, составляет 14 тыс. оснований. В середине самого большого экзона 26 имеется глутаминовый кодон CAA, который в результате редактирования мРНК превращается в нонсенс-кодон UAA, что приводит к образованию более короткой мРНК длиной в 7 тыс. оснований. В результате трансляции редактированной мРНК образуется укороченный AроB48. Полноразмерный белок Аро100 синтезируется в клетках печени и вовлечен в транспорт эндогенно синтезированных триглицеридов и холестерола, АроВ48 образуется в клетках кишечника и задействован в транспорте жиров, поступающих с пищей и всасывающихся в кишечнике.
Помимо указанных механизмов регуляции избирательной экспрессии генов на уровне процессинга РНК есть еще один - регуляция транспорта мРНК из ядра. Например, у млекопитающих лишь около 5% синтезированной РНК покидает ядро и транспортируется в цитоплазму. Достоверно неизвестно, каким образом происходит отбор конкретных РНК, подлежащих участию в трансляции. Однако изучение геномов различных млекопитающих позволяет указать один из механизмов такого отбора. Суть его заключается в том, что в молекулу РНК, служащую матрицей для синтеза белка, вносятся изменения, делающие молекулу «бессмысленной», и тогда она уничтожается. Обнаружено, что подобные изменения вносятся в ультраконсервативные участки ДНК, которые абсолютно идентичны у разных групп млекопитающих, в частности, у человека и мыши. Установлено, что таким образом происходит регуляция экспрессии генов, белки которых принимают участие в осуществлении альтернативного сплайсинга.
Следующий уровень регуляции избирательной экспрессии генов - уровень трансляции. Даже при одинаковом наборе готовых к трансляции мРНК клетки могут различаться между собой по времени начала и по скорости трансляции. Регуляция трансляции включает в себя репрессию-дерепрессию собственно трансляции, а также стабилизацию-дестабилизацию мРНК (см. п. 2.4.5.6). Наиболее общий пример регуляции такого рода - блокирование трансляции заготовленных в оогенезе материнских мРНК вплоть до активации яйцеклетки. Инактивацию и стабилизацию материнских мРНК в ооцитах обеспечивают определенные белки, например FRGY2 у шпорцевой лягушки или p50 у кроликов, участвующие в запасании мРНК в составе рибонуклеопротеиновых частиц - информосом.
В дальнейшем по ходу дробления материнская мРНК вступает в трансляцию также не сразу повсеместно, а по определенной пространственно-временной программе. Устранение блока трансляции на временно инактивированных мРНК после активации яйцеклетки достигается в том числе добавлением большого количества адениловых групп на 3'-конце молекул. Полиаденилирование происходит у многих эукариотических организмов и является крайне консервативным механизмом регуляции функционирования мРНК на ранних стадиях развития. Полиаденилирование осуществляется поли(А)-полимеразами (PAP), одна из которых находится в ядре, а другая локализована в цито плазме.
Существенные задержки в начале трансляции уже заготовленных мРНК отмечены также при дифференцировке эритроидных, спермато-генных и других специализированных типов клеток.
Изменение скорости трансляции наблюдается, например, в ходе дифференцировки клеток хрусталика куриного зародыша: на 6-е сутки эмбрионального развития на 1 молекулу мРНК синтезируется в 5 раз больше соответствующего белка (α-кристаллина), чем на 19-е сутки.
Недавно был открыт новый способ регуляции на уровне трансляции, основанный на так называемой РНК-интерференции. Это явление впервые было обнаружено у круглого червя Caenorhabditis elegans, а к настоящему времени выявлено у многих эукариотических организмов, в том числе у человека. В клетке на требуемых стадиях развития благодаря активности определенных генов и последующему процессингу возникают короткие молекулы РНК (размером 21-28 нуклеотидов). Они связываются с комплементарными участками транслируемых мРНК, что приводит к подавлению синтеза белка молекулы, а иногда и к деградации активных мРНК. Возможность РНК-интерференции установлена в цитоплазме яйцеклетки насекомых.
После завершения трансляции осуществляется посттрансляционная регуляция экспрессии гена. Вновь синтезированный полипептид, прежде чем стать функционально активным, проходит многочисленные превращения, например отщепление фрагментов, различные химические модификации, например добавление фосфатных или углеводных групп (ацетилирование, фосфорилирование и гликозилирование), изменение третичной структуры, образование в ряде случаев четвертичной структуры из нескольких субъединиц, наконец, так называемую «адресацию» - перемещение к месту окончательного функционирования.
Время и место посттрансляционных превращений, как правило, строго определены. Временная задержка посттрансляционных модификаций может быть достаточно большой. Например, фермент тирози-наза появляется у зародышей амфибий еще в раннем эмбриогенезе, но переходит в активную форму лишь после вылупления зародыша. Роль посттрансляционных модификаций в регуляции клеточной дифферен-цировки изучена еще далеко не достаточно, но предполагают, что она весьма значительна.
Таким образом, представленные в данном разделе механизмы регуляции избирательной экспрессии генов объясняют, каким образом в ходе индивидуального развития может осуществляться процесс диффе-ренцировки различных типов клеток. Следует иметь в виду, что избирательной экспрессии подвергаются, как правило, не отдельные гены, а целые группы (блоки) генов, обеспечивающие специфическую диффе-ренцировку клеток конкретного типа. После активации блока генов их экспрессия поддерживается на определенном уровне. Этим может быть объяснена высокая устойчивость дифференцированного состояния многих типов клеток.
Однако формирование целостного организма в ходе индивидуального развития не сводится только к приобретению специфичности конкретными группами клеток, поэтому применительно к многоклеточному организму клеточная дифференцировка неотрывна от пространственно-временных аспектов и, следовательно, от еще более высоких уровней ее регуляции, нежели уровни регуляции биосинтеза белка на клеточном уровне. На организменном уровне регуляция дифференцировки осуществляется благодаря системным механизмам развития или механизмам интеграции, реализация которых обеспечивает целостность и интегрированность организма на всех стадиях его онтогенеза, начиная с момента образования зиготы. Подробно эти механизмы рассматриваются в следующих разделах.
Зрелая яйцеклетка, которую Т.Х. Морган справедливо считал самой дифференцированной клеткой в организме, представляет собой мозаичную, высокогетерогенную систему. Один из процессов, приводящий к гетерогенности яйцеклетки - овоплазматическая сегрегация.
Неравномерное распределение компонентов цитоплазмы в яйцеклетке можно обнаружить уже на стадии созревания. Как было показано в п. 7.2, овоплазматическую сегрегацию сопровождает поляризация яйцеклетки. Остановимся более подробно на процессах, приводящих к овоплазматической сегрегации и роли последней в дифференцировке клеток развивающегося зародыша.
Формирование сегрегации цитоплазмы подробно изучено в созревающем яйце дрозофилы. Неоднородность цитоплазмы яйцеклетки возникает в том числе вследствие неравного положения ее полюсов среди клеток материнского организма. К переднему полюсу яйцеклетки примыкают фолликулярные и питающие клетки, которые продуцируют мРНК для белка bicoid. Эта мРНК транспортируется в яйцеклетку, и еще до оплодотворения устанавливается градиент ее концентрации с максимумом на переднем конце яйцеклетки, что обусловливает в дальнейшем развитии формирование головных структур зародыша из этой части яйца. Будущий задний полюс соседствует с фолликулярными клетками, которые доставляют в эту область яйца мРНК, считанную с гена nanos, детерминирующую образование заднего конца зародыша (рис. 8.26). Таким образом, еще в неоплодотворенной яйцеклетке дрозофилы формируется передне-задняя ось будущего организма. Аналогично задается и дорсо-вентральная ось. Вещества, имеющие четкий градиент распределения, принято называть морфогенами. Следует обратить внимание, что градиенты морфогенов в яйцеклетке дрозофилы создаются благодаря активности в окружающих яйцо фолликулярных и питающих клетках генов материнского организма. Они получили название - «гены с материнским эффектом».

Рис. 8.26. Распределение морфогенов по продольной оси яйца дрозофилы
Следует обратить внимание, что большинство запасенных в яйцеклетке мРНК первоначально находится в неактивном состоянии в комплексе с белком в виде информосом. Такие неактивные мРНК могут быть распределены в цитоплазме достаточно равномерно. Создание их градиентов осуществляется благодаря их неравномерной активации. Механизмы активации могут быть различными. Так, например, установлено, что фиксация мРНК на цитоплазматическом матриксе яйцеклетки (локализация мРНК в определенных зонах яйца) приводит к их активации. Показано, что в дальнейшем транслируются только локализованные мРНК, а нелокализованные разрушаются.
Другие механизмы избирательной трансляции запасенных мРНК наблюдаются на следующих этапах формирования овоплазматической сегрегации и связаны с перемещениями цитоплазмы вследствие оплодотворения.
Проникновение сперматозоида в яйцеклетку в момент оплодотворения и последующее движение его пронуклеуса приводит к усилению овоплазматической сегрегации. В яйце наблюдаются сложные перемещения цитоплазмы и ее функциональная перестройка. В результате она становится еще более неоднородной. Этот процесс хорошо заметен в тех случаях, когда разные участки цитоплазмы содержат гранулы веществ разной окраски (желток, темный пигмент и др.).
Хорошим примером может служить яйцо асцидии. Серая цитоплазма центральной части яйцеклетки окружена кортикальным слоем, содержащим желтые липидные включения. На анимальном полюсе располагается светлая цитоплазма с ядерным материалом. Сразу после оплодотворения цитоплазма яйца перемещается, и кортикальный ее слой формирует желтый серп, расположенный между экватором и вегетативным полюсом (рис. 8.27).
Перемещения цитоплазмы вследствие оплодотворения хорошо заметны и в яйцеклетке амфибий. В ней слой темного пигмента меланина первоначально покрывает все анимальное полушарие. После проникновения сперматозоида поверхностный - кортикальный - слой цитоплазмы толщиной в несколько микрометров поворачивается примерно на 30о относительно внутренней массы желтка в направлении, которое зависит от места проникновения сперматозоида. В результате этого у некоторых амфибий против места проникновения спермия появляется серповидная слабопигментированная область, названная серым серпом. В ней позже, в ходе гаструляции, возникает дорзальная губа бластопора. Вследствие всех указанных перемещений цитоплазмы формируются оси зародыша. Сторона, где формируется серый серп, становится дорзаль-ной, а противоположная, где наблюдается внедрение сперматозоида - вентральной. Анимально-вегетативная ось соответствует головно-хво-стовой оси будущего зародыша (см. рис. 7.3).
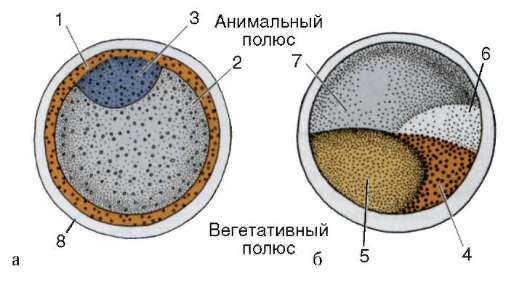
Рис. 8.27. Сегрегация цитоплазмы в яйце асцидии Styelapartita: а - до оплодотворения; б - после оплодотворения; 1 - кортикальная цитоплазма с желтыми липидными включениями; 2 - цитоплазма, содержащая желток; 3 - светлая цитоплазма с ядром ооцита; 4 - желтый серп; 5 - цитоплазма с желтком; 6 - серый серп; 7 - анимальная светлая цитоплазма; 8 – хорион
Перемещения цитоплазмы вследствие проникновения спермия в яйцеклетку - еще один механизм, приводящий к избирательной трансляции запасенных в яйцеклетке мРНК. В частности, благодаря в том числе и этому механизму осуществляется детерминирование дорсо-вентральной оси зародыша. В оогенезе у амфибий на вегетативном полюсе яйца запасаются мРНК для белка Xwnt11. После оплодотворения и поворота цитоплазмы часть этой мРНК перемещается по стороне, противоположной внедрению сперматозоида, в направлении анималь-ного полюса. В области серого серпа происходит полиаденилирование молекул мРНК Xwnt11, что приводит к их активации и последующей трансляции. В результате только в этой области яйца образуется соответствующий белок - один из основных дорсализующих факторов. Остальная мРНК для Xwnt11 в вегетативном полушарии, по-видимому,остается репрессированной. Выполненные исследования установили, что именно поворот цитоплазмы является механизмом, запускающим трансляцию мРНК Xwnt11 через полиаденилирование.
Анализ результатов многих проведенных экспериментов позволил сделать вывод, что в создании сегрегации цитоплазмы яйцеклетки ведущая роль принадлежит цитоскелету. Так, транспорт мРНК, поступающих из окружающих клеток и синтезированных в самой яйцеклетке, к месту их локализации в цитоплазме осуществляется на большие расстояния по микротрубочкам, а на малые - по микрофиламентам. Считают, что местом локализации в клетке морфогенетических детерминант может быть кортикальный слой или цитоскелет клетки. Предполагают также, что и перемещения цитоплазмы яйца, наблюдаемые после оплодотворения, определяются цитоскелетом. В частности, в этом процессе возможно значительное участие центриоли сперматозоида и отходящих от нее микротрубочек. С помощью нарушающего сборку микротрубочек колхицина удается подавить транспорт и активацию мРНК, перемещения цитоплазмы и ооплазматическую сегрегацию в целом.
В ходе дробления разные участки цитоплазмы зиготы, содержащие специфический набор веществ, попадают в разные бластомеры. Экспериментами с микроинъекциями коллоидных частиц золота показано, что при дроблении цитоплазма яйцеклетки распределяется между бла-стомерами, не перемешиваясь. Различия в характере цитоплазмы могут служить регулятором считывания информации с разных генов в разных бластомерах и тем самым влиять на ход их дифференцировки. Показано, что цитоплазматические факторы белковой природы проникают в ядро бластомера и путем избирательной активации или инактивации конкретных генов определяют характер считываемой информации. Полагают, что таким способом морфогенетические детерминанты, содержащиеся в отдельных участках цитоплазмы, жестко, порой необратимо контролируют предопределенность (детерминированность) данного бластомера к образованию клеток определенного типа.
Такое жесткое предопределение судьбы бластомеров наблюдается у оболочечников, к которым относятся и асцидии. Неоднократно экспериментально доказано, что у этих животных каждый бластомер ответственен за образование специфического набора тканей личинки, при этом каждая клетка дифференцируется автономно, независимо от окружающих ее клеток. Тунг и другие исследователи пересаживали в цитоплазму бластомера, лишенного генетического материала, ядро из другого бла-стомера. Показано, что дальнейшее развитие клетки-реципиента идет по пути того бластомера, чья цитоплазма ему досталась. Обнаружено также, что удаление у оболочечников каких-либо бластомеров приводит к отсутствию у личинки как раз тех структур, которые в норме из них формируются, а изоляция определенных групп клеток зародыша приводит к формированию из них характерных структур вне связи с другими клетками. Так, в серии экспериментов на 8-клеточном зародыше оболо-чечников показана способность только одной пары бластомеров, содержащих фермент ацетилхолинэстеразу, давать начало мышечной ткани.
У асцидий после оплодотворения по-разному окрашенные области цитоплазмы яйца распределяются по разным бластомерам, детерминируя их дальнейшую судьбу. Клетки бластулы, унаследовавшие цитоплазму желтого серпа, дают начало мышечным клеткам, цитоплазму серого экваториального серпа - образуют хорду и нервную трубку, анимальную цитоплазму - становятся эпидермисом личинки, содержащие желток вегетативной области - формируют в ходе развития кишку (см. рис. 8.27).
Жесткая детерминация судьбы бластомеров, определяемая составом веществ попавшего туда участка цитоплазмы яйца, обнаружена и у ряда других животных, например гребневиков, круглых и кольчатых червей, моллюсков. Тип развития этих животных, дифференцировка клеток которых определяется очень рано в развитии благодаря прежде всего ово-плазматической сегрегации, назван мозаичным.
Помимо овоплазматической сегрегации в определении судьбы бла-стомеров на самых ранних этапах развития может принимать участие и другой системный механизм - межклеточные взаимодействия. В этом случае развитие бластомеров в большей степени зависит от их взаимодействий с соседними клетками, межклеточным матриксом, которые определяются положением этих бластомеров в зародыше. Подобный тип развития, наблюдаемый у иглокожих и позвоночных, назван регуляционным.
Следует, однако, иметь в виду, что в развитии и мозаичных, и регуляционных зародышей участвуют оба механизма, однако степень их влияния значительно разнится, и основную роль играет один из них. Так, локализация специфических белков или м-РНК в определенных областях зиготы не ограничена мозаичными зародышами. Обнаружено, что анимальные и вегетативные области яиц амфибий, имеющих регуляционный тип развития, содержат уникальные мРНК. Кроме того, в цитоплазме вентральной области зиготы лягушки была выявлена так называемая половая детерминанта. Клетки, получающие при дроблении цитоплазму с данным веществом, становятся предшественниками половых клеток, и их потомки дают начало гаметам. Показано, что у зародышей ряда других животных, раннее развитие которых является в основном регуляционным и определяется межклеточными взаимодействиями, обнаруживаются подобные половые детерминанты. Содержащие их бластомеры в ходе дальнейшего развития дают начало предшественникам гамет и мигрируют в закладку гонад.
Помимо овоплазматической сегрегации, гетерогенность яйцеклетки определяется также неоднородностью организации ее плазмалем-мы. Так, для овулировавших яйцеклеток млекопитающих характерна своеобразная организация цитоскелета, что в свою очередь приводит к мозаичной организации плазматической мембраны. Основная часть мембраны яйцеклетки образует микроворсинки, и лишь примерно от одной десятой до одной пятой общей поверхности яйцеклетки мыши представлено районом, в котором нет микроворсинок. Под плазма-леммой в этой области яйцеклетки располагается густая сеть микро-филаментов, а глубже находится мейотическое веретено метафазы II. У других млекопитающих район, не имеющий микроворсинок, также соответствует той области цитоплазмы, где располагается мейотическое веретено.
При оплодотворении спермий контактирует с мембраной яйцеклетки в любом месте, богатом микроворсинками. После этого микроворсинки исчезают, генетический материал спермия попадает в цитоплазму яйцеклетки, а часть мембраны спермия встраивается в месте его проникновения в мембрану яйцеклетки. В результате возникает разнородность плазматической мембраны, которая отражается на нескольких делениях дробления. Первые два бластомера имеют одинаковый размер, но не вполне одинаковые характеристики. Во время интерфазы синтез рибосомальной РНК в ядрышках этих бластомеров происходит в разное время, продолжительность фазы репликации ДНК у них также неодинакова. Один из бластомеров содержит в плазмалемме антигены, а в цитоплазме - структурные компоненты хвоста спермия. Предполагается, что именно этот бластомер вступает во второе деление дробления на 20-60 мин раньше другого. Мембранные антигены спермия сохраняются в плазмалеммах у потомков этого бластомера еще на протяжении нескольких делений. Установлено, что потомки именно этого бластомера, который на 2-клеточной стадии делится первым, с большей вероятностью дадут начало развитию внутренней клеточной массы бла-стоцисты, тогда как потомки запаздывающего при делении бластомера с большей вероятностью станут источником для формирования внезаро-дышевых частей эмбриона.
Таким образом, гетерогенность яйцеклетки не только определяет последующую дифференцировку клеток зародыша, но и обеспечивает развитие зародыша как единой системы.
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 2440; Нарушение авторских прав?; Мы поможем в написании вашей работы!