КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
I-й этап аэробного распада углеводов
|
|
|
|
АЭРОБНЫЙ РАСПАД УГЛЕВОДОВ
Это главный путь распада углеводов до конечных продуктов, а для клеток некоторых тканей является единственно возможным. В среднем аэробному распаду подвергается 70-75% глюкозы, поступающей в клетку, и для многих тканей этот самый протяженный путь распада углеводов является главным источником получения энергии в клетке. Получение энергии в виде АТФ и является главным биологическим значением аэробного распада углеводов.
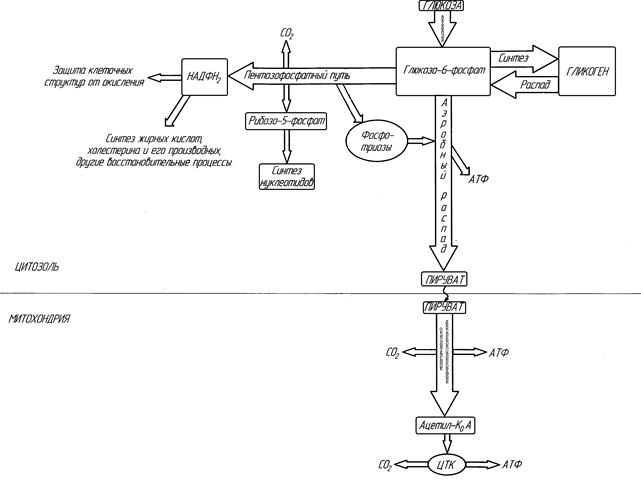
Рисунок ХХ. Главные пути метаболизма углеводов (схема).
Аэробный путь распада углеводов условно можно разделить на три этапа.
Протекает в цитоплазме, включает в себя 10 реакций и заканчивается образованием двух молекул пирувата (пировиноградной кислоты).
Условно I-й этап аэробного распада можно разделить на 2 части. Первая часть представляет из себя постепенную активацию молекулы гексозы, завершающуюся ее распадом на две триозы. Такая активация протекает в 2 стадии, первая из которых описана выше (гексокиназная реакция). В ходе вторичной активации в состав молекулы углевода включается еще один остаток фосфорной кислоты, в результате чего образуется бисфосфатная форма гексозы - фруктозо-1,6-бисфосфат:
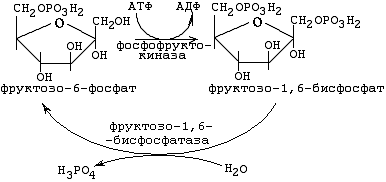
Рисунок ХХ. Реакция, катализируемая фосфофруктокиназой и ее обходной обратный путь
Реакция необратима, сопровождается затратой одной молекулы АТФ и катализируется ферментом фосфофруктокиназой. Фосфофруктокиназа - это один из ключевых ферментов аэробного пути распада углеводов. Максимальная скорость работы фосфофруктокиназы больше, чем Vmax гексокиназы. Поэтому, когда в клетку в период интенсивного энергопотребления поступает много глюкозы, скорость всего аэробного пути распада углеводов (как, впрочем, и остальных путей метаболизма глюкозы) ограничивается именно гексокиназой.
Однако в условиях низкого энергопотребления (например, в покоящейся мышце) накапливаются АТФ и цитрат, избыток которых сильно ингибирует фосфофруктокиназу. В такой ситуации лимитирующим ферментом аэробного пути распада углеводов вместо гексокиназы становится именно фосфофруктокиназа. Из-за ее угнетения в клетке накапливаются фруктозо-6-фосфат и его предшественник глюкозо-6-фосфат. Избыток последнего, в свою очередь, ингибирует гексокиназу, уменьшая использование глюкозы клеткой в целом, и одновременно активирует гликогенсинтетазу, запуская синтез гликогена.
В условиях дефицита энергии, когда нет избытка АТФ и цитрата, а существует избыток АДФ, то последний активирует фосфофруктокиназу, и тогда скорость всего аэробного пути лимитируется опять гексокиназой.
В результате фосфофруктокиназной реакции молекула гексозы дестабилизируется (ослабляется) настолько, что образующийся в ней фруктозо-1,6-бисфосфат легко распадается на 2 фосфотриозы. Эти вещества – фосфоглицериновый альдегид (сокращенно – ФГА) и фосфодиоксиацетон (сокращенно – ФДА) являются изомерами и способны к взаимному превращению. Так завершается начальная часть первого этапа аэробного распада распада углеводов, в ходе которой клетка не только не получает энергии, но вынуждена вкладывать ее в виде двух молекул АТФ. Но, как мы увидим в дальнейшем, эти энергетические затраты многократно окупаются в последующих реакциях аэробного распада, и клетка получает прибыль в виде большого количества АТФ, несравнимо большую по сравнению с затратами в начальной части первого этапа процесса.
В следующую реакцию первого этапа аэробного пути распада углеводов вступает только ФГА. В результате уменьшается его концентрация, и постепенно весь ФДА переходит в ФГА. Поэтому во вторую часть первого этапа аэробного распада вступают сразу две молекулы ФГА, образовавшиеся из одной молекулы гексозы.
Вторая часть 1-го этапа начинается с окисления ФГА до ФГК (фосфоглицериновой кислоты) в 6-й реакции.
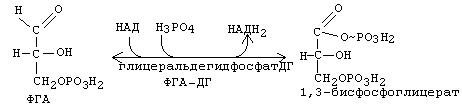
Рисунок ХХ. Реакция окисления фосфоглицеринового альдегида.
Эта обратимая реакция катализируется ферментом глицеральдегидфосфатдегидрогеназой. Эта никотинамидная дегидрогеназа способна передавать отщепляемый водород со своего кофермента по системе транспортных челночных механизмов в митохондрию на полную дыхательную цепь, и затем на кислород воздуха, с образованием 2,5 молекул АТФ (путем окислительного фосфорилирования).
Для первого этапа аэробного распада указанная реакция является единственно возможной точкой сопряжения с работой митохондриальных дыхательных цепей посредством транспортных челночных механизмов. Поэтому для первого этапа аэробного распада углеводов характерна лишь относительная независимость от наличия кислорода в клетке.
Поскольку окислению подвергаются сразу две молекулы ФГА, образовавшиеся из одной гексозы, то общий энергетический итог этой реакции - образование 5 АТФ в расчете на одну молекулу глюкозы, либо на один глюкозный фрагмент молекулы гликогена. Энергии, которая выделяется при этом окислении, хватает и на то, чтобы одновременно обеспечить присоединение фосфата к альдегидной группе. Присоединяется фосфат макроэргической связью. В результате образуется 1,3-дифосфоглицериновая кислота (1,3-бисфосфоглицерат). Макроэргическая связь реализуется в следующей реакции: происходит субстратное фосфорилирование, и клетка получает еще 2 молекулы АТФ. И, наконец, в последней, 10-й, необратимой реакции первого этапа аэробного распада, также происходит субстратное фосфорилирование с образованием двух АТФ в расчете на одну молекулу гексозы.
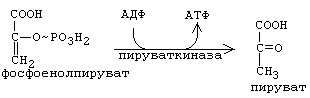
Рисунок ХХ. Пируваткиназная реакция, завершающая I-й этап аэробного распада углеводов
Образующаяся пировиноградная кислота - это конечный продукт первого этапа аэробного распада углеводов.
В целом, энергетику первого этапа аэробного распада глюкозы можно описать следующим образом:
1. Затрата 2 АТФ на активацию глюкозы (гексокиназная и фосфофруктокиназная реакции).
2. Образование 5 АТФ путем окислительного фосфорилирования при окислении двух молекул ФГА с участием челночных транспортных механизмов.
3. Две реакции субстратного фосфорилирования с образованием 2 АТФ в каждой из них (всего 4 АТФ).
Таким образом, с учетом затрат, общий энергетический выход для первого этапа аэробного распада в расчете на одну молекулу глюкозы составляет 7 молекул АТФ. В случае же распада гликогена отщепляемый глюкозный фрагмент сразу же образуется в виде глюкозо-6-фосфата, и затраты АТФ на гексокиназную реакцию в этом случае не происходит. Поэтому в случае распада глюкозного фрагмента гликогена в ходе первого этапа аэробного распада клетка получает на одну молекулу АТФ больше, чем в случае распада глюкозы – всего 8 молекул АТФ.
Пируват, образующийся в ходе первого этапа аэробного распада, транспортируется в митохондрию, где и подвергается последующим превращениям.
Второй и третий этапы аэробного распада углеводов являются исключительно аэробными. Они протекают в митохондриях с обязательным участием кислорода, и посредством многих реакций жестко связаны с работой дыхательных цепей митохондриального окисления. Основное количество молекул АТФ, получаемое клеткой при аэробном распаде углеводов, образуется именно в ходе II-го и III-го этапов процесса.
II-й этап аэробного распада углеводов – «окислительное декарбоксилирование пировиноградной кислоты»
II-й этап катализируется мультиферментным комплексом окислительного декарбоксилирования α-кетокислот (подробнее этот комплекс и катализируемые им реакции описаны в главе «Митохондриальное окисление»). В ходе II этапа от молекулы пирувата отнимаются два атома водорода с одновременным декарбоксилированием, и образуется продукт – Ацетил-кофермент А (активная форма уксусной кислоты). Отнятый водород передается по цепи митохондриального окисления на кислород воздуха с образованием Н2О и 2,5 молекул АТФ. Поскольку из молекулы глюкозы образуется 2 молекулы пирувата, то II-й этап приносит клетке 5 АТФ в расчете на одну молекулу глюкозы.
III-й этап аэробного распада углеводов – цикл трикарбоновых кислот
(ЦТК, цикл Кребса, лимоннокислый цикл)
В III этап аэробного распада вступает молекула АцетилКоА - продукт II-го этапа, взаимодействуя при этом со щавелевоуксусной кислотой. Химизм реакций ЦТК подробно описан в главе «Митохондриальное окисление». В этом циклическом процессе, включающем в себя 8 реакций, протекающих в матриксе митохондрий, ацетильные остатки полностью расщепляются до конечных продуктов метаболизма: СО2 и Н2О. При этом образуется 10 АТФ в расчете на одну молекулу АцетилКоА, вступившую в цикл и полностью «сгоревшую» в нем. Если оценить энергетический итог процесса в расчете на 1 молекулу глюкозы или 1 глюкозный фрагмент гликогена, то на III-м этапе аэробного распада углеводов образуется 20 АТФ.
Если к этим 20 АТФ добавить те молекулы АТФ, которые образуются в расчете на одну молекулу гексозы, вступившей в аэробный распад, на I-м и II-м этапах – 7 АТФ (8 АТФ - для глюкозного остатка гликогена) и 5 АТФ соответственно, то суммарный энергетический итог всего аэробного пути распада углеводов составит 32 АТФ на каждую молекулу глюкозы, расщепленную до СО2 и Н2О или 33 АТФ в расчете на каждую молекулу глюкозного остатка гликогена.
ИТОГОВОЕ УРАВНЕНИЕ АЭРОБНОГО РАСПАДА ГЛЮКОЗЫ:

В образующихся молекулах АТФ заключено 65% энергии, которая выделилась бы при сжигании глюкозы на воздухе. Это доказывает очень высокую эффективность работы аэробного пути распада углеводов.
Цикл трикарбоновых кислот содержит еще одно ключевое звено аэробного пути распада углеводов. Ключевое звено на 3-м этапе является необходимым, поскольку АцетилКоА, вступающий в ЦТК, образуется не только из углеводов, но и из жиров и аминокислот. Поэтому ЦТК - это конечный "котел" для сжигания ацетильных остатков, образующихся из веществ разных классов, и объединяет все метаболиты, образующиеся при распаде углеводов, жиров и белков.
Ключевые ферменты ЦТК цитратсинтетаза и изоцитратдегидрогеназа угнетаются избытком АТФ и избытком НАДН2, а изоцитратдегидрогеназа активируется избытком АДФ.
Особенность регуляции ЦТК заключается в том, что АТФ ингибирует эти ферменты по-разному: изоцитратдегидрогеназа ингибируется АТФ намного сильнее, чем цитратсинтаза. Поэтому при избытке АТФ накапливаются промежуточные продукты: цитрат и изоцитрат. В этих условиях цитрат может выходить в цитоплазму по градиенту концентраций, что важно, прежде всего, для обмена липидов.
II-й и III-й этапы аэробного пути распада углеводов протекают в митохондриях, а I-й - в цитоплазме. Таким образом, I-й этап отделен от II-го и III-го митохондриальной мембраной и, в отличие от II-го и III-го этапов, возможно его обращение.
7 реакций I-го этапа обратимы полностью. Для остальных 3-х необратимых стадий существуют обходные обратные реакции.
Для гексокиназной реакции (1-я стадия) обходной обратный путь катализируется глюкозо-6-фосфатазой (см. рис. ХХ).
Для фосфофруктокиназной реакции (3-я стадия) обходной обратный путь катализируется фруктозо-1,6-бисфосфатазой (см. рис. ХХ).
Третьим необратимым звеном I-го этапа является его последняя, 10-я стадия. В прямом направлении эта реакция катализируется пируваткиназой (см. рис. ХХ), а обходной обратный путь протекает в 2 стадии. В 1-й реакции СО2 присоединяется к ПВК с образованием щавелево-уксусной кислоты (ЩУК). Эта реакция синтеза катализируется пируваткарбоксилазой и требует затраты 1-й молекулы АТФ. В состав кофермента пируваткарбоксилазы входит биотин – витамин H. ЩУК поступает в цитоплазму (с участием челночных механизмов), где превращается в ФЕП с затратой 1-й молекулы ГТФ, что эквивалентно затрате 1-й АТФ.
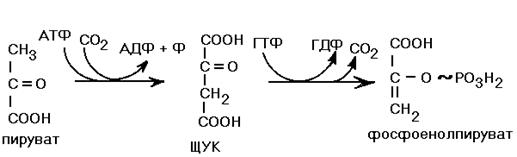
Рисунок ХХ. Реакция, катализируемая пируваткарбоксилазой (обходной обратный путь для пируваткиназной реакции).
С учетом обращения двух реакций субстратного фосфорилирования и затрат НАДН2 на обращение реакции окисления фосфоглицеринового альдегида (см. рис. ХХ) путь синтеза глюкозы из пирувата требует расходования 12 АТФ в расчете на каждую синтезированную молекулу глюкозы. Это подтверждает правило, что на любой биосинтез определенного метаболита требуется энергии больше, чем образуется при распаде этого же метаболита (в данном случае необходимо 12 АТФ для синтеза глюкозы против 7 АТФ, образующихся при ее распаде).
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 2211; Нарушение авторских прав?; Мы поможем в написании вашей работы!