КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Зависимость скорости реакции от рН 4 страница
|
|
|
|
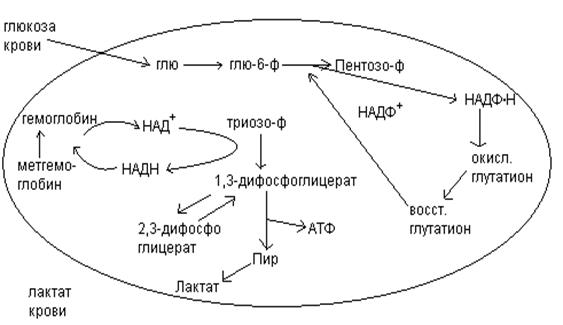
Обмен углеводов в эритроцитах.
Эритроциты не содержат ядра, митохондрий. В эритроците не идут реакции цикла лимонной кислоты, в них нет ферментов дыхательной цепи. Парадоксальным является тот факт, что эритроцит, перенося кислород для тканей, сам его не использует и получает энергию за счет аэробных процессов.
Основным процессом в эритроцитах, который дает энергию, является анаэробный гликолиз. При расщеплении фру-6-фф образуется НАДН, необходимый для восстановления избытка метгемоглобина (окисленной формы гемоглобина, не связывающей О2).
Побочным продуктом гликолиза в эритроцитах является 2,3-дифосфоглицерат. 2,3-дифосфоглицерат связывается с гемоглобином, уменьшает его сродство к О2 и, облегчает освобождение кислорода в тканях.
Фосфоглюконатный путь в норме составляет лишь небольшую долю в катаболизме глюкозы. В условиях повышенной потребности в НАДФ.Н этот процесс активизируется. НАДФ.Н необходим для того, чтобы поддерживать внутриклеточный восстановитель, глутатион, в его восстановленной SH-форме. Воздействие агентов, ускоряющих окисление глутатиона в S-S-форму, активирует реакции пентофосфотного пути, которые обеспечивают образование восстановленных эквивалентов в форме НАДФ.Н.
Обмен углеводов в эритроцитах.
Регуляция обмена углеводов.
Соотношение между процессами катаболизма и анаболизма глюкозы в клетках печени находятся под контролем целого ряда факторов регуляции:
1. Концентрация метаболитов и глюкозы.
2. Воздействие гормонов.
Внутриклеточные рецепторы.
Субстраты и промежуточные продукты метаболических реакций играют важную роль в регуляции соотношения между гликолизом и глюконеогенезом.
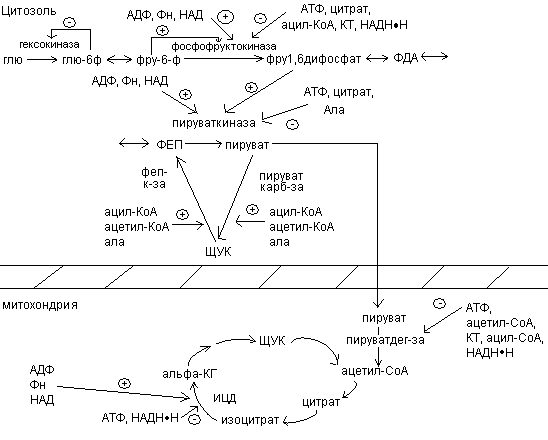
Рассмотрим, как регулируется само вступление остатков глюкозы на путь гликолиза. Вовлечение глюкозных остатков в процесс гликолиза обеспечивает важная реакция и эта реакция контролируется регуляторным ферментом. Реакция катализируется гексокиназой и активность этого фермента ингибируется глюкозо-6-фосфатом. Когда концентрация глюкозо-6-фосфата в клетке сильно возрастает, т.е. когда он образуется быстрее, чем потребляется, наступает ингибирование – гексокиназа под действием глюкозо-6-фосфата выключается и дальнейшего фосфорилирования глюкозы не происходит до тех пор, пока избыток глюкозо-6-фосфата не будет использован.

 Глю
Глю  Глю-6-ф
Глю-6-ф
 В печени преобладает другой фермент – глюкокиназа, которая не ингибируется глюкозо-6-фосфатом. Поэтому в печени, способной хранить большие количества гликогена, избыточная глюкоза крови может фосфорилироваться в глюкозо-6-фосфат, который через глюкозо-1-фосфат превращается в гликоген.
В печени преобладает другой фермент – глюкокиназа, которая не ингибируется глюкозо-6-фосфатом. Поэтому в печени, способной хранить большие количества гликогена, избыточная глюкоза крови может фосфорилироваться в глюкозо-6-фосфат, который через глюкозо-1-фосфат превращается в гликоген.
Кроме гексокиназной реакции в гликолизе имеются еще два главных регулируемых этапа: это фосфофруктокиназная и пируваткиназная реакция.
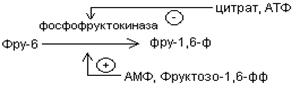
В скелетных мышцах активность фосфофруктокиназы определяется концентрациями субстратов этого фермента (АТФ и фруктозо-6-фосфата) и его продуктов (АДФ и фруктозо-1,6-дифосфата), а также цитрат.
Главными отрицательными модуляторами фосфофруктокиназой являются АТФ и цитрат.
Главными положительными модуляторами являются АМФ и фруктозо-1,6-дифосфат.
Третьим регулируемым этапом гликолиза является пируваткиназная реакция.

Активность пируваткиназы ингибируется АТФ и ацетил-КоА, а также жирные кислоты.
| Реакция | Фермент | Ингибитор | Активатор |
| Глю Глю-6-ф
| Глю-6-ф | ||
Фру-6-ф  Фру-1,6-дф Фру-1,6-дф
| АТФ, цитрат, ацил-КоА, кетоновые тела, НАДН+Н+ | Фру-1,6-дф, АДФ, НАД | |
ФЕП 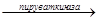 Пир Пир
| АТФ, ацетил-КоА, жирные к-ты, аланин | АДФ, Фн, НАД, фру-1,6-дф | |
Пир 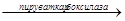 ЩУК ЩУК
| Ацил-КоА, Ацетил-КоА, Аланин | ||
ЩУК 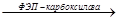 ФЭП ФЭП
| Ацил-КоА, Ацетил-КоА, Аланин |
Регуляция гликолиза.
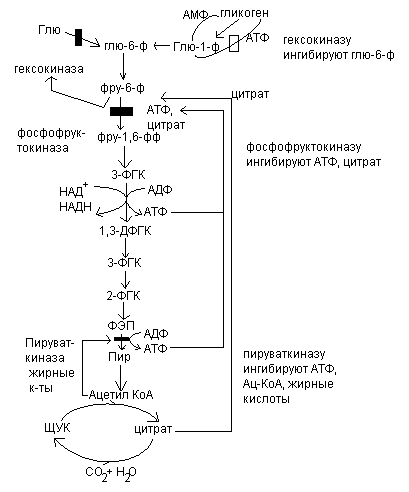
Три стадии катаболизма углеводов обеспечивают получение энергии: гликолиз, цикл лимонной кислоты и окислительное фосфорилирование.
При уменьшении содержания АТФ и Рн, это приводит к возрастанию скорости переноса электронов и окислительного фосфорилирования. Одновременно повышается скорость окисления пирувата через цикл лимонной кислоты, т.е. усиливается приток электронов в дыхательную цепь. Это приводит к увеличению скорости гликолиза, тем самым обеспечивается образование пирувата. Затем наступает момент, когда отношение АТФ/АДФ+Рн возвращается к обычному высокому уровню. Теперь перенос электронов и окислительное фосфорилирование замедляются. Цикл лимонной кислоты замедляется, т.к. АТФ и цитрат являются ингибитором фосфофруктокиназы и пируваткиназы.
Регуляция пируватдегидрогеназного комплекса.
Превращение пирувата в Ацетил-КоА происходит при участии пируватдегидрогеназного комплекса, содержащего 5 коферментов: НАД, ФАД, НSКоА, ТДФ, липоевую кислоту. В основе процесса лежит реакция окислительного декарбоксилированя.
Образование ацетил-КоА из пирувата – это ключевой необратимый этап метаболизма, потому что животные неспособны к превращению ацетил-КоА в глюкозу.
Окислительной декарбоксилирование пирувата в ацетил-КоА создает возможность превращения атомов глюкозы по 2 путям: 1) окисление до СО2 в ЦТК с одновременным регенерированием энергии; 2) включение в липиды. Это позволяет считать, что активность пируватдегидрогеназного комплекса должна строго регулироваться.
Активность ПДК (пируватдегидрогеназного комплекса) регулируется 3 путями:
1. Ингибирование продуктами реакции. Ацетил-КоА и НАДН ингибируют превращение
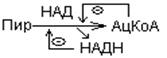
2. Регуляция нуклеотидами по принципу обратной связи.
Пируватдегидрогеназный комплекс ингибируется ГТФ и активируется АМФ.
Активность комплекса снижается, когда клетка богата легкодоступной энергией.
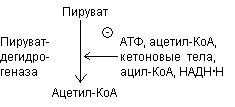
3. Регуляция путем ковалентной модификации. Комплекс теряет ферментативную активность, когда специфический остаток серина пируватдегидрогеназного комплекса фосфорилируется АТФ. Фосфорилирование усиливается при высоких соотношениях АТФ\АДФ, ацетил-КоА\СоА, НАДН\НАД+ и ингибируется пируватом.
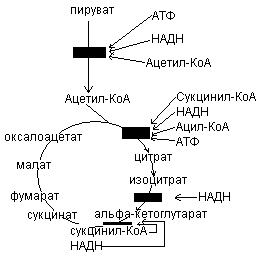
Регуляция ЦТК.
Скорость функционирования ЦТК зависит от потребности клетки в АТФ. Важной регуляторной реакцией цикла является синтез цитрата из ацетил-КоА и оксалоацетата под действием цитратсинтетазы. АТФ – аллостерический ингибитор цитратсинтетазы. 1) Ац-КоА + ЩУК 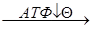 цитрат
цитрат
Вторая регуляторная реакция – это реакция, катализируемая изоцитрат-дегидрогеназой. Фермент аллостерически активируется АДФ, НАДН ингибирует изоцитрат-дегидрогеназу:
2) Изоцитрат 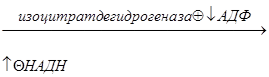
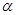 -оксоглутарат
-оксоглутарат
3) Третьей регуляторной реакцией является реакция, катализируемая -кетоглутаратдегидрогеназой:
-кетоглутарат 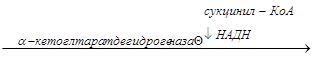 сукцинил-КоА
сукцинил-КоА
оксоглутарат
Регуляция цикла лимонной кислоты.
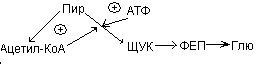
Активация пируваткарбоксилазы.
Активность пируваткарбоксилазы зависит от присутствия ацетил-КоА.
Пир может превращаться в ацетил-КоА, а также подвергаться карбоксилированию с образование оксалоацетата или ЩУК. Высокое содержание ацетил-КоА приводит к образованию количества оксалоацетата. Если имеет место избыток АТФ, то оксалоацетат потребляется в процессе глюконеогенеза:
Ацетил-КоА 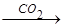 ЩУК
ЩУК 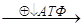 ФЕП
ФЕП  Глю
Глю
В условиях недостатка АТФ оксалоацетат включается в ЦТК, конденсируясь с ацетил-КоА.
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 422; Нарушение авторских прав?; Мы поможем в написании вашей работы!