КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Нуклеосомы обычно упаковываются вместе, образуя при этом упорядоченные структуры высшего порядка
|
|
|
|
9-10
9.1.18. Некоторые нуклеосомы расположены на ДНК неслучайным образом [16]
Опыты, проведенные in vitro с изолированным хроматином, дают основание полагать, что при физиологических условиях гистоновые октамеры остаются зафиксированными в одном положении, поскольку их тесная связь с нуклеиновой кислотой препятствует перемещению по спирали ДНК. Остается открытым вопрос, случайно или нет расположены эти октамеры на ДНК (случайность следует понимать так, что в одной клетке определенная последовательность ДНК плотно обвита вокруг гистонового октамера, а в другой - та же последовательность отделяет друг от друга бусины нуклеосом).
Для того чтобы определить положение нуклеосом в клетках, нужно обработать их ферментом или реагентом, вносящим разрывы в ДНК, а затем изучить защищенные от воздействия участки методом, аналогичным футпринтингу ДНК (см. разд. 4.6.6). Хотя большинство нуклеосом, по-
видимому, расположены случайным образом, известны поразительные примеры из неслучайного расположения. Так, у дрожжей Saccharomy-ces
cerevisiae 15 нуклеосом строго фиксированным образом окружают ДНК центромеры (последовательность CEN) (разд. 13.5.3). Единственный сайт локализации имеет и нуклеосома, связанная с очень маленьким по размеру геном 5S-pPHK. Известно также, что по крайней мере одна нуклеосома расположена перед точкой начала синтеза РНК β-глобина, Чем же объясняется такое неслучайное расположение нуклеосом? Показано, что в некоторых случаях (например, для нуклеосом, связанных с генами 5S-pPHK) смесь четырех очищенных гистонов, составляющих нуклеосому, in vitro образует ее точно на том же месте, где она расположена in vivo. Возможно, причина заключается в том, что нуклеосомы стремятся связаться таким образом, чтобы максимально заполнить богатую AT малую бороздку ДНК. Такое предпочтение вызвано тем, что двойную спираль ДНК трудно уложить двумя плотными витками вокруг гистонового октамера, и для этого требуется значительное уплотнение на малой бороздке спирали ДНК (рис. 9-24). Как установлено на примере белка репрессора бактериофага (см. рис. 9-17), кластер, состоящий из двух или трех пар AT и расположенный в малой бороздке, облегчает возникновение такого уплотнения. На характер расположения нуклеосом должны влиять и другие неизвестные пока свойства последовательности ДНК.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
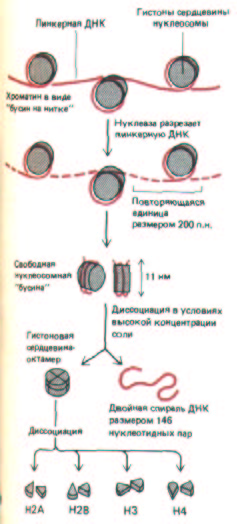
Рис. 9-23. Строение нуклеосом. Нуклеосомные частицы состоят из двух полных витков ДНК (83 нуклеотидных пары на виток), закрученных вокруг кора, представляющего собой гистоновый октамер, и соединяются между собой линкерной ДНК. Нуклеосом-ная частица выделена из хроматина путем ограниченного гидролиза линкерных участков ДНК микрококковой нуклеазой. В каждой нуклеосомнои частице фрагмент двойной спирали ДНК, имеющий в длину 146 пар оснований, закручен вокруг гистонового кора. Этот белковый кор содержит по две молекулы каждого из гистонов Н2А, Н2В, НЗ и Н4. Полипептидные цепи гистонов насчитывают от 102 до 135 аминокислотных остатков, а общий вес октамера составляет приблизительно 100000 Да. В деконденсированной форме хроматина каждая «бусина» связана с соседней частицей нитевидным участком линкерной ДНК.
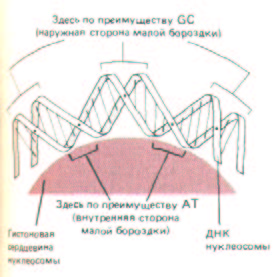
Рис. 9-24. Изгиб ДНК на нуклеосоме. Спираль ДНК делает два оборота вокруг гистонового октамера, причем на каждый виток приходится по 83 нуклеотидные пары. Соотношение ДНК и белка на схеме приближено к реальному, чтобы показать сдавливание малой бороздки на внутренней части витка. Как указывалось выше (см. рис. 9-17), в узкой малой бороздке расположены главным образом АТ-пары оснований.
9.1.19. Определенные сайты на хромосомах не содержат нуклеосом [17]
На некоторых участках ДНК нукдеосомы отсутствуют, несмотря на то, что длина этих участков составляет сотни нуклеотидных пар.
Такие области можно выявить, обработав ядро клетки следовыми количествами дезоксирибонуклеазы (ДНКаза 1). Использование минимальных концентраций фермента обеспечивает разрушение длинных областей безнуклеосомной ДНК, при этом короткие участки линкерной ДНК, расположенной между нуклеосомами, останутся целыми. Хроматин, обработанный таким образом, расщепляется преимущественно по участкам, которые, по-видимому, не содержат нуклеосом. Обычно такие сайты отстоят друг от друга на расстояние нескольких тысяч нуклеотидных пар.
Первое доказательство биологической значимости сайтов, сверхчувствительных к нуклеазе, было получено в экспериментах с вирусом SV40. Его хромосома помимо кольцевой ДНК содержит гистоны, продуцируемые клеткой-хозяин ом. В составе этой хромосомы имеется участок длиной 300 нуклеотидных пар, который свободен от нуклеосом и быстро разрушается под воздействием ДНКазы!. Этот участок расположен очень близко от последовательностей ДНК, с которых начинается как репликация ДНК вируса, так и синтез его РНК. Здесь же локализуются и несколько сайт-специфических ДНК-связывающих белков, которые защищают лишь небольшой участок этой молекулы, по-
видимому, совершенно лишенный нуклеосом, от нуклеазной деградации. Аналогичным образом, многие участки хроматина в клетке, обладающие гиперчувствительностью к ДНКазе, расположены в регуляторных областях генов (рис. 9-25); в клетках, где эти гены активны, таких сайтов больше, нежели в других клетках. Полагают, что за удаление нуклеосом ответственны сайт-специфические ДНК-связывающие белки, которые принимают участие в регуляции эукариотических генов (см. рис. 9-27).
[18]
В живых клетках хроматин, вероятно, редко имеет вид растянутых «бус на нитке». Напротив, нуклеосомы связаны друг с другом и образуют регулярные структуры, в которых ДНК еще больше конденсирована. Электронномикроскопический анализ лизированных прямо на сеточке ядер показывает, что большая часть хроматина имеет форму фибрилл диаметром около 30 нм (рис. 9-22, А). Один из возможных способов упаковки нуклеосом в фибрилле хроматина диаметром 30 нм приведен на рис. 9-26. Эта модель представляет идеальную структуру. В действи-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
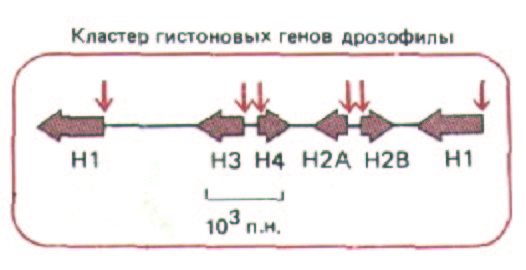
Рис. 9-25, Расположение сайтов, сверхчувствительных к нуклеазе (цветные стрелки), в регуляторних областях активных генов. Хотя такие участки хроматина обычно расположены на 5'-конце гена, как показано на этом рисунке на примере кластера генов, кодирующих гистоны (Н., Н2А, Н2В, НЗ и Н4) у дрозофилы, эти сайты могут находиться и в других областях (см. рис. 10-40, А).
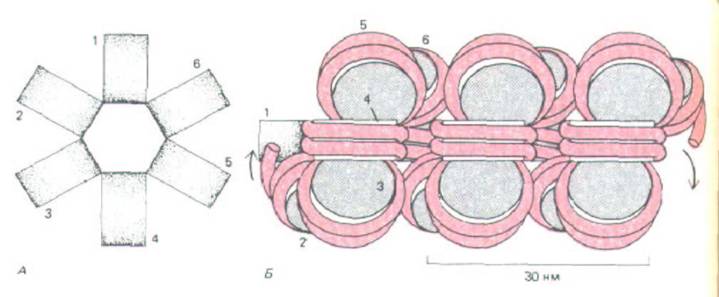
Рис. 9-26. Модель, предложенная для объяснения упаковки нуклеосомной нити в составе 30 нм-фибриллы, наблюдаемой под электронным микроскопом (см. рис. 9-22, А). А. Вид сверху. Б. Вид сбоку. При таком типе упаковки на нуклеосому приходится одна молекула гистона Н1 (не указано). Хотя место прикрепления гистона Н1 к нуклеосоме определено, расположение молекул Н1 на этой фибрилле неизвестно (см. также рис. 9-27).
тельности и разброс длин линкерных участков (что связано с определенным расположением нуклеосом), и наличие образовавшихся случайным образом безнуклеосомных последовательностей придают различным участкам 30 нм-фибриллы различные свойства (рис. 9-27).
Если бы хроматин типичной хромосомы человека существовал в виде фибриллы диаметром 30 нм, то в растянутом состоянии он имел бы длину 0,1 см, т.е. в 100 раз превышал бы размеры ядра. Микроскопический анализ интактных хромосом дает основание полагать, что внутри клеток происходит дальнейшая упаковка фибрилл диаметром 30 нм, при этом образуются нити хроматина толщиной 100 нм. Как расположены нуклеосомы в такой структуре, неясно.
9.1.21. Гистоновые белки Н1 помогают соединять нуклеосомы [19]
В клетках млекопитающих имеется приблизительно шесть близкородственных вариантов (подтипов) гистонов Н1, которые несколько отличаются друг от друга по аминокислотной последовательности. По-видимому, эти молекулы ответственны за упаковку нуклеосом в фибриллу диаметром 30 нм. Молекулы Н1 имеют эволюционно консервативную глобулярную центральную область, соединенную с выступающими аминоконцевыми и карбоксильными «ручками», аминокислотная последовательность которых эволюционирует быстрее. Каждая молекула Н1
связывается своей глобулярной частью с уникальным сайтом на нуклеосоме, а «ручка», как полагают, захватывают область шире и контактируют с другими сайтами, расположенными на гистонах, входящих
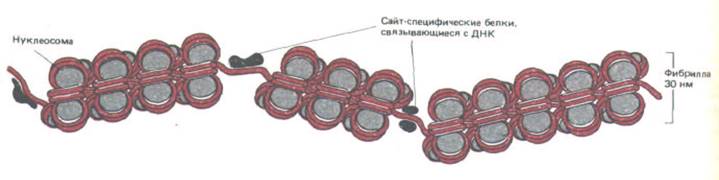
Рис. 9-27. Схема, иллюстрирующая прерывание правильной нуклеосом-нон структуры хроматина короткими областями, в которых ДНК необычно чувствительна к обработке ДНКазой.. В каждом из этих сайтов, сверхчувствительных к нуклеазе, нуклеосомы на ДНК, вероятно, замещены одним или несколькими сайт-специфическими ДНК-связывающими белками.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
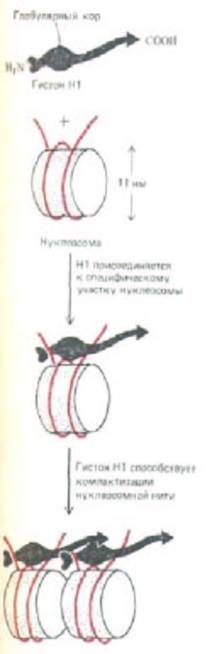
Рис. 9-28. Схема, показывающая, каким образом гистон Н1 (220 аминокислот) мог бы обеспечить контакт соседних нуклеосом.
Глобулярная часть Н1. связывается с каждой нуклеосомой вблизи сайта, в котором спираль ДНК входит и выходит с гистонового октамера. В присутствии гистона Н1 два полных витка ДНК (166 нуклеотидных пар) защищены от действия микрококковой нуклеазы (см. рис. 9-23). Однако до сих пор неизвестны ни трехмерная структура гистона Н1., ни точные области взаимодействия выступающих аминоконцевых и карбоксиконцевых плечей этого гистона с нуклеосомой.
в состав соседних нуклеосом. При этом нуклеосомы стягиваются вместе, образуя регулярную повторяющуюся структуру (рис. 9-28). Согласно разным гипотезам, молекулы Н1 располагаются либо внутри хроматиновой 30 нм-фибриллы, либо снаружи.
Гистоновый октамер, образующий сердцевину каждой нуклеосомы, представляет собой симметричную структуру, а единственная молекула гистона Н1, связывающаяся с каждой нуклеосомой, симметрией не обладает. Таким образом, связывание молекул Н1 с хроматином создает локальную полярность (рис. 9-29).
Опыты, проведенные in vitro, показывают, что при связывании гистона H1 с ДНК к ней присоединяется сразу по восемь или более белковых молекул, что может служить примером кооперативного связывания. Весьма вероятно, что хроматин организован именно за счет кооперативных взаимодействий такого типа, а при их нарушении белками-регуляторами происходит локальная деконденсация хроматина (что и имеет место в области активных генов). Образовавшиеся области «активного хроматина», по-видимому, обладают необычайно низкой способностью связывать гистон Н1. В этом отношении ограниченная область хроматина может напоминать крошечный кристалл, который способен в ходе процессов, приводящих к активации гена, менять свою конформацию по типу все или ничего (рис. 9-30).
9.1.22. Нуклеосомы не мешают синтезу РНК [20]
Результаты биохимических исследований свидетельствуют о том, что при транскрипции большая часть ДНК остается связанной с нуклеосомами; электронномикроскопическое изучение препаратов распластанного по специальной методике хроматина обычно выявляет одинаковое расположение нуклеосом как в транскрибируемых, так и в нетранскриби-
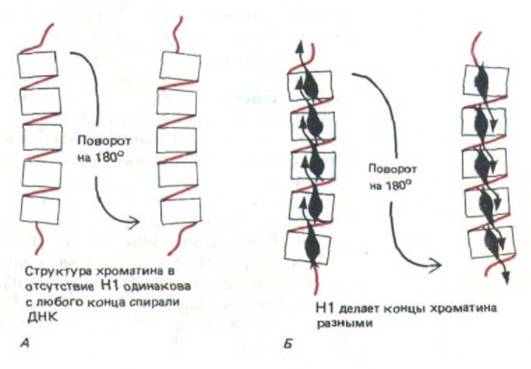
Рис. 9-29. Полярность хроматина обусловлена связыванием его с гистоном Н1. А. В отсутствие гистона Н1 хроматин не обладает полярностью, так как каждая нуклеосома симметрична. Б. В присутствии Н. хроматин становится полярным. Остается неизвестным, для чего нужна такого рода полярность.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
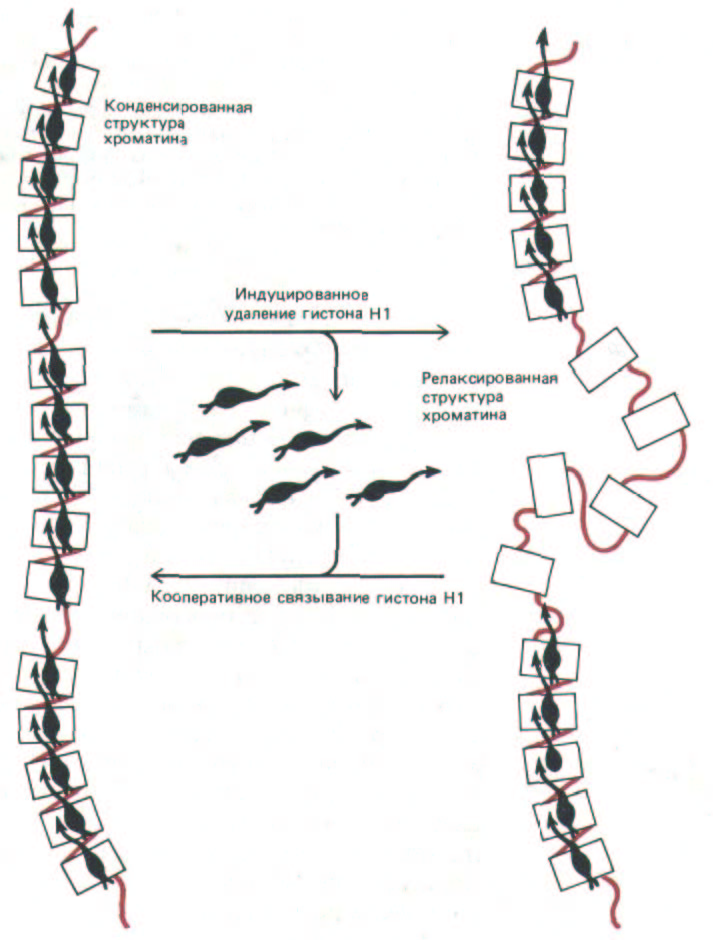
Рис. 9-30. Отдельные участки хроматина могут вести себя как структурные единицы вследствие кооперативного характера упаковки нуклеосом с помощью гистона Н1. Приведенная схема показывает момент внезапной деконденсации такой единицы (фибриллы 30 им или более компактной структуры), вызванной внешним регуляторним сигналом. Деконденсация хроматина подобного типа может сопутствовать активации гена (см. рис. 9-50).
руемых областях (рис. 9-31). По-видимому, гистоновый октамер так прочно связан с ДНК, что остается на ней постоянно. Тем не менее трудно себе представить, как РНК-полимераза транскрибирует ДНК, связанную с нуклеосомой, не внося никаких временных изменений в строение нуклеосом.
По мере прохождения РНК-полимеразы, ДНК, входящая в состав нуклеосомы, вероятно, раскручивается с гистонового октамера, не освобождая его полностью. Возможно, вся нуклеосома временно приоткрывается, разделяя гистоновый октамер на две половинки. Не исключено также, что октамер остается интактным, но как бы «покачивается», давая пройти полимеразе (рис. 9-32). Очевидная трудность подобных маневров может служить объяснением столь высокой консервативности, которая характерна для аминокислотных последовательностей гистонов. Если бы нуклеосома представляла собой лишь устройство для наматывания ДНК, было бы логично ожидать большей вариабельности аминокислотной последовательности у нуклеосомных гистонов.
Как уже обсуждалось выше, большая часть нуклеосом клетки упакована в хроматиновую фибриллу 30 нм, которая затем подвергается дальнейшей конденсации. Трудно себе представить, что хроматин, находящийся в таком состоянии, транскрибируется РНК-полимеразой, и при этом упаковка нуклеосом, входящих в его состав, не подвергается значительным изменениям (рис. 9-33). Некоторые эксперименты дают основание предполагать, что отчасти такое изменение структуры имеет место (см. рис. 9-50).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
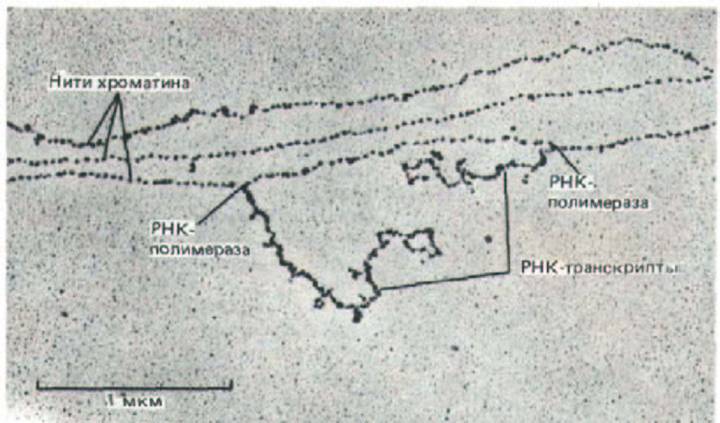
Рис. 9-31. Участок хроматина в форме нуклеосомной нити. Показаны три нити хроматина, на одной из которых две молекулы РНК-
полимеразы транскрибируют ДНК. Большая часть хроматина в ядре высших эукариот не содержит активных генов, и, следовательно, свободна от РНК-транскриптов. Следует отметить, что нуклеосомы имеются как в транскрибируемых, так и в нетранскрибируемых областях, и что они связаны с ДНК непосредственно перед и сразу же за движущимися молекулами РНК-полимераз. (С любезного разрешения Victoria Foe.).
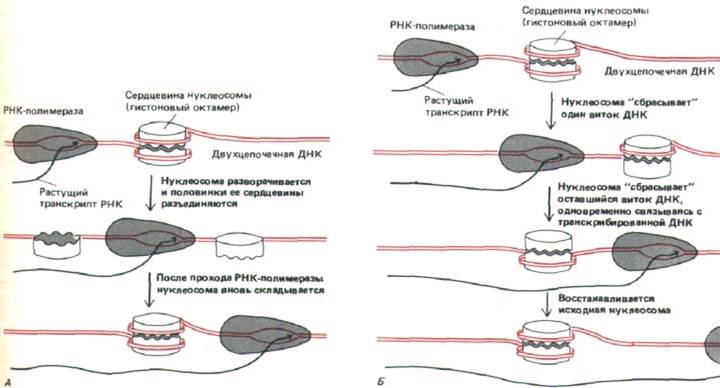
Рис. 9-32. Две возможные модели, объясняющие, как РНК-полимераза может транскрибировать хроматин, не вытесняя с него нуклеосомы. А. Транскрипция через временно приоткрытые полинуклеосомы. Б. Транскрипция через целый гистоновый октамер.
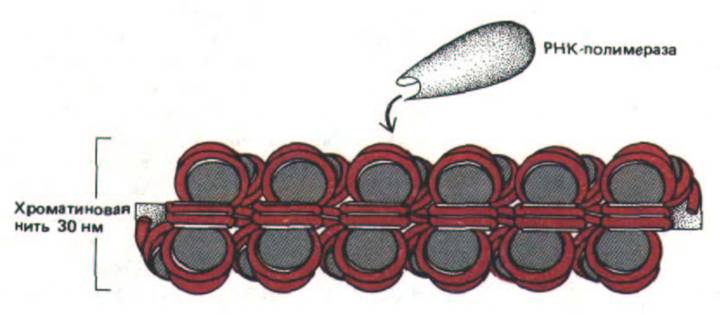
Рис. 9-33. Схема, представляющая молекулу РНК-лолимеразы, которая приближается к 30 нм-фибрилле хроматина. Число изображенных нуклеосом соответствует примерно 7000 парам оснований ДНК, что сопоставимо с геном среднего размера у человека (табл. 9-.).
Вся эта ДНК, не соскальзывая с гистоновых октамеров хроматина, каким-то образом должна оказаться доступной для действия РНК-полимеразы.
Очевидно, что для этого необходимо развернуть хроматин.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
Заключение
Ген - это последовательность нуклеотидов, представляющая собой единицу активности для образования молекулы РНК. Хромосома
состоит из одной-единственной невероятно длинной молекулы ДНК, содержащей множество генов. В молекуле хромосомной ДНК имеются и
другие типы нуклеотидных последовательностей, необходимых для ее функционирования: сайт инициации репликации и теломера (они
обеспечивают репликацию молекулы ДНК), а также центромера (она служит для прикрепления ДНК к митотическому веретену). Гаплоидный
геном человека содержит 3 х 109 нуклеотидных пар, которые распределены между 22 различающимися аутосомами и 2 половыми хромосомами.
По-видимому, лишь несколько процентов этой ДНК кодируют белки.
ДНК эукариот тесно связана с большим количеством гистонов, которые служат для образования множества повторяющихся частиц, содержащих белки и ДНК и называемых нуклеосомами. Нуклеосомы обычно упакованы вместе в регулярную структуру — фибриллу, имеющую
диаметр 30 нм. Однако в областях ДНК, содержащих гены, гистоновый октамер, образующий каждую нуклеосому, должен конкурировать с
разнообразными сайт-специфическими белками за участки связывания на ДНК. Такие участки, на которых нуклеосома замещена ДНК-
связывающими белками, обычно выявляются как области более активной транскрипции ДНК.
В эукариотической клетке имеются сотни самых разнообразных сайт-специфических ДНК-связывающих белков. Каждый из них узнает
короткую последовательность ДНК по водородным связям с парами оснований и по форме спирали. Эти белки при образовании белковых
комплексов на определенных участках ДНК вступают друг с другом в кооперативное или конкурирующее взаимодействие.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 3489; Нарушение авторских прав?; Мы поможем в написании вашей работы!