КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Репликация хромосом
|
|
|
|
9-15
9-14
9.2.8. В транскрипционно активных областях хроматин менее конденсирован [28]
При обработке ядер клеток позвоночных ДНКазой I (той самой, которая при более низких концентрациях выявляет сверхчувствительные к нуклеазе области, лишенные нуклеосом) было показано, что опреде-
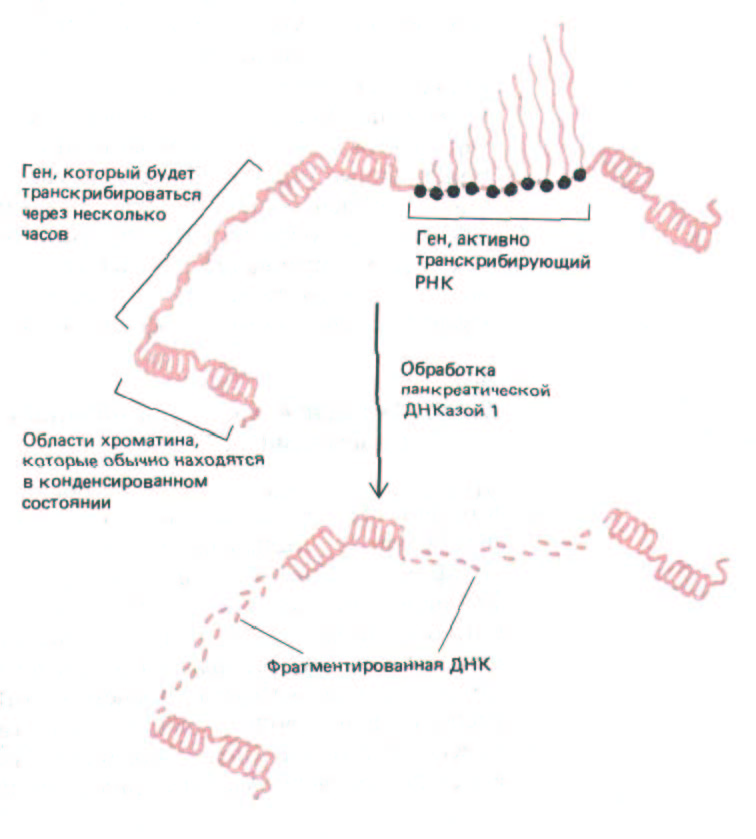
Рис. 9-51. Обработка хроматина панкреатической ДНКазой I. Вначале фермент разрезает сайты, обладающие повышенной чувствительностью к нуклеазе (не показано), затем деградации подвергается последовательность ДНК, в которую входят активно транскрибируемые и потенциально активные гены.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.

Рис. 9-52. Электронная микрофотография расправленного хроматина из клеток эмбриона насекомого (Oncopeltus). Видны значительные изменения в структуре хроматина двух тандемно расположенных генов рРНК. По-видимому, у этих эмбрионов синтезу рРНК предшествует переход хроматина из состояния «бусины на нитке» к гладкой форме. Возможно, гладкий хроматин представляет собой область, где ДНК «соскочила» с нук-леосом в ходе подготовки к транскрипции. Следует отметить, что структура хроматина, входящего в состав гена, расположенного левее от транскрибируемой области, подверглась изменениям, хотя в данный момент он не транскрибируется. (По V. Е. Foe, Cold Spring Harbor Symp. Quant. Biol. 42: 723-740, 1978.)
ленная часть хроматина, содержащего активные гены, находится в де-конденсированном состоянии. Этот вывод подтверждает результаты, полученные на политенных хромосомах насекомых. При определенной концентрации ДНКазы в первую очередь подвергается деградации около 10% генома позвоночных. В разных клетках одного и того же организма нуклеаза расщепляет различные последовательности ДНК, которые соответствуют различным типам продуцируемых этими клетками РНК. С помощью гибридизации со специфическими ДНК-зондами удалось выяснить, что деградировавшие последовательности относятся главным образом к тем областям генома, которые в данном типе клеток активно транскрибируются. Интересно, что даже гены, транскрибирующиеся в каждом поколении клеток лишь несколько раз, обладают чувствительностью к нуклеазе. Этот факт говорит о том, что сверхчувствительным к нуклеазе участок становится скорее всего вследствие особого состояния хроматина, а не собственно транскрипции (рис. 9-51). Хроматин в таком чувствительном к нуклеазе состоянии часто называют активным
хроматином. Полагают, что нуклеосомы, входящие в его состав, упакованы менее плотно. Даже если нуклеазной обработке подвергаются митотические хромосомы, расщепленным оказывается преимущественно активный хроматин. По-видимому, и в митозе, несмотря на чрезвычайно плотную упаковку генома, между активным и неактивным хроматином сохраняются некоторые различия.
Электронномикроскопический анализ, как правило, не выявляет никаких изменений в строении нуклеосом, входящих в состав активного хроматина. Однако есть и поразительное исключение: у некоторых зародышей насекомых в области активных генов рРНК можно заметить протяженные участки хроматина, как будто лишенного нуклеосом (рис. 9-52). Согласно одной из гипотез, в таких участках каждая нуклеосома открывается, образуя развернутую структуру. Вероятно, при этом происходит раскручивание двух витков спирали ДНК, которые в обычном состоянии намотаны на нуклеосому. По-видимому, эти структурные изменения способствуют эффективности транскрипции.
9.2.9. Активный хроматин обладает особыми биохимическими свойствами [29]
Для того чтобы разобраться в строении хроматина, сконденсированного в разной степени, необходимо выделить и охарактеризовать хромосомные белки, присущие каждому из этих состояний. В отношении активного хроматина на этом пути были достигнуты определенные успехи. Установлено, что 1) гистон Н1 не очень тесно связан по крайней мере с некоторой частью активного хроматина; 2) четыре гистона, образующие нуклеосому, содержатся в обычных количествах, но характеризуются необычно высоким уровнем ацетилирования лизиновых остатков, расположенных вблизи аминоконца этих белков. Ацетильные группы присоединяются к ним с помощью фермента гистоновой ацетилазы
и удаляются гистоновой деацетилазой, причем каждая ацетильная груп-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
па существует в среднем около 10 минут; 3) нуклеосомы в активном хроматине селективно связывают два близкородственных небольших хромосомных белка HMG14 и HMG17. Так как эти белки присутствуют только в активном хроматине, их количество строго соответствует тому, которое требуется для связывания примерно одной из каждой 10 нуклеосом. Аминокислотные последовательности обоих белков крайне консервативны, что свидетельствует о важности их функции; 4) у некоторых организмов, например у Tetrahymena, активный хроматин в значительной мере обогащен минорной формой гистона Н2А. Подобная разновидность гистона найдена также у дрозофилы, курицы и человека.
Любое из описанных свойств может играть важную роль в развора-чивании хроматина и таким образом способствовать синтезу РНК, однако для подтверждения этой гипотезы требуется проведение прямых экспериментов. Дальнейшие биохимические исследования можно проводить с использованием новых методов очистки активного хроматина на основе селективного связывания с аффинным матриксом, который содержит антитела, специфические либо к белку HMG, либо к ацетилиро-ванному лизину. В гл. 10 обсуждаются механизмы контроля образования активного хроматина.
9.2.10. Гетерохроматин сильно конденсирован и транскрипционно неактивен [30]
Доказательства существования второй необычной формы хроматина были получены в 1930 г., когда при наблюдении в световом микроскопе интерфазного ядра клетки высших эукариот была обнаружена особая конденсированная форма хроматина, названная гетерохроматином (менее конденсированная часть хроматина была названа эухроматином). Гетерохроматин в интерфазе остается необычайно компактным, сохраняя такую структуру, которую большая часть хроматина приобретает только в митозе. Позднее обнаружилось, что как и митотический хроматин, Гетерохроматин транскрипционно неактивен (рис. 9-53). Полагают, что в большинстве клеток около 90% хроматина транскрипционно неактивно. Хотя относительная устойчивость такого неактивного хроматина к обработке нуклеазой свидетельствует в пользу того, что этот хроматин более конденсирован, чем 10% в транскрипционно
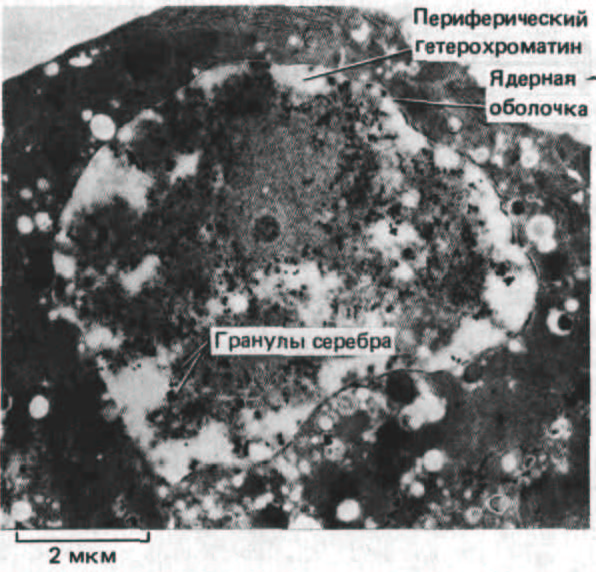
Рис. 9-53. Радиоавтограф тонкого среза ядра клетки, подвергнутой импульсному мечению [3Н]-уридином для выявления областей активного синтеза РНК (гранулы серебра). Светлые участки - области гетерохроматина, который обычно концентрируется вдоль внутренней поверхности ядерной оболочки. Светлая окраска участков гетерохроматина обусловлена особенностями метода приготовления препарата. Синтез РНК наблюдается главным образом в эухроматино-вых участках, окружающих области гетерохроматина. (С любезного разрешения Stan Fakan.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
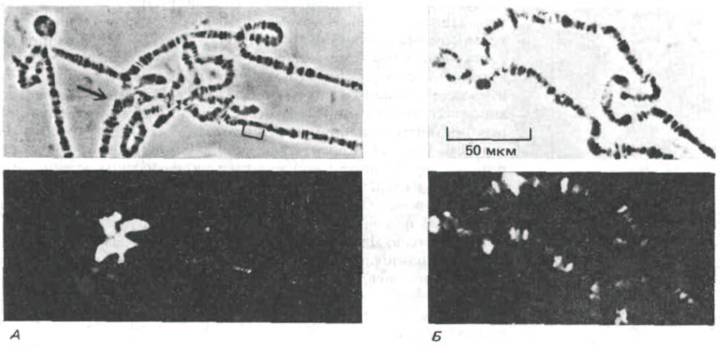
Рис. 9-54. Участок политенной хромосомы дрозофилы в фазово-контрастном (вверху) и флуоресцентном (внизу) микроскопе после иммунофлуоресцентного мечения гетерохроматина (слева) и активного хроматина (справа). А. Окрашивание моноклональными антителами, специфичными к белкам, содержащимся в гетерохроматиновых областях. Окрашены хромоцентр (указан стрелкой) и несколько отдельных дисков.
Б. Окрашивание моноклональными антителами, специфичными к белкам, содержащимся в активном хроматине. Окрашивается часть дисков. (Л-по Т.С. James and S.C.R. Elgin, Моl. Cell Biol. 6: 3862 3872, 1986; Я-по G.C. Howard, S.M Abmayr, Z.A. Shinefeld, V.Z. Sato and S.C.R. Elgin, J. Cell Biol.
88: 219-225, 1981. © 1981 Rockefeller University Press.)
активных областях, лишь часть его (возможно от 10 до 20%) обладает сильно конденсированной конформацией, известной под названием гетерохроматин. Таким образом, гетерохроматин, вероятно, представляет собой особый класс транскрипционно инертного хроматина, несущего определенные функции. У млекопитающих и многих других высших эукариот ДНК, окружающая каждую центромеру, состоит из относительно простых повторяющихся нуклеотидных последовательностей; именно такие «сателлитные ДНК» составляют основную часть гетерохроматина этих организмов.
У дрозофилы в виде гетерохроматина организованы как хроматин центромеры, так и случайно разбросанные короткие участки.
Подобные гетерохроматиновые области недореплицируются на ранних стадиях синтеза ДНК, т. е. в тот период, когда происходит образование политенной хромосомы. Таким образом, эти последовательности в политенных хромосомах представлены в относительно меньших количествах.
Биохимические особенности этого типа гетерохроматина можно изучать на молекулярном уровне, проводя связывание с антителами, выявляющими белки хромосом, которые присутствуют только в гетерохроматине (рис. 9-54). Перспективным для биохимического анализа гетерохроматина представляется и клонирование генов, кодирующих специфические для него белки.
Заключение
Все хромосомы во время митоза подвергаются значительной конденсации. Специальное окрашивание выявляет на митотических
хромосомах полосы, благодаря которым можно совершенно точно идентифицировать каждую хромосому. Эти полосы, насчитывающие
миллионы нуклеотидных пар ДНК, выявляют в хромосоме значительную гетерогенность, природа которой пока еще неясна.
Во время интерфазы хромосомы обычно подвергаются деконденсации в такой степени, что их структура становится
трудноразличимой. Замечательным исключением являются специализированные хромосомы типа ламповых щеток в ооцитах позвоночных и
политенные хромосомы гигантских секреторных клеток насекомых. Изучение этих двух разновидностей интерфазных хромосом показало, что
каждая длинная молекула ДНК, входящая в состав хромосомы, состоит из большого числа
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
дискретных, по-разному упакованных доменов. Основная характерная черта этих хромосом — наличие значительного количества петель
хроматина, большая часть которых содержит от 20 000 до 100 000 нуклеотидных пар. Как в хромосомах типа ламповых щеток, так и в
политенных хромосомах наименее конденсированными являются области, наиболее активно синтезирующие РНК. Судя по чувствительности к
ДНКазе, лишь 10% ДНК находится в относительно деконденсированном состоянии и транскрибируется. Такой «активный» хроматин имеет
иные биохимические свойства, нежели те, которыми обладают более конденсированные участки.
Перед каждым делением клетка должна синтезировать копии всех своих хромосом. Таким образом, делению клетки предшествует ее переход из состояния интерфазы (фазы G1) в фазу синтеза ДНК (S-фаза). В типичной клетке высших эукариот S-фаза длится 8 часов. После ее окончания каждая хромосома представлена двумя копиями, которые продолжают оставаться соединенными в области центромер до наступления М-фазы. (см. рис. 9-35). Для удвоения хромосомы необходима репликация ее ДНК и последующая сборка на этой молекуле хромосомных белков, образующих хроматин. В гл. 5 мы обсуждали ферменты, участвующие в репликации ДНК, и строение репликационной вилки, обеспечивающей синтез (см. рис. 5-39). Переход клетки в S-фазу будет рассмотрен в гл. 13 как часть более общей проблемы контроля клеточного цикла. В данном разделе мы изложим принципы механизма репликации эукариотической хромосомы, укажем время, необходимое для этого, и, кроме того, проанализируем взаимосвязь процесса репликации и структуры хромосомы.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 1423; Нарушение авторских прав?; Мы поможем в написании вашей работы!