КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Комбинации нескольких регуляторных белков, контролирующих активность генов, могут определять
|
|
|
|
развитие многих типов клеток [4]
Принципиальная схема комбинационной регуляции активвости гена приведена на рис. 10-7, где каждый пронумерованный элемент соответствует отдельному белку-регулятору. На этой чисто гипотетичной схеме одна материнская клетка дает начало двум типам клеток А и В, различающимся лишь тем, что в одной из них происходит синтез регуляторного белка 1, а в другой-нет. В ходе дальнейшего развития этих клеток в некоторых из них появляются регуляторные белки 2 и 3, а затем 4 и 5. В результате формируются 8 типов клеток (они обозначены буквами от G до N), характеризующиеся различными сочетаниями пяти регуляторных белков. Добавление к приведенной схеме еще двух белков-регуляторов (6 и 7)
повлечет за собой образование на следующей стадии 16 типов клеток. Еще 10 подобных стадий в сумме дадут 10000 различных клеточных типов-и все это за счет взаимодействия только 25 регуляторных белков.
Таким образом, комбинационная регуляция активности гена весьма эффективна: малое число регулирующих элементов может детерминировать нормирование сложных биологических объектов В гл. 16 будет показано, как эта система функционирует в ходе развития
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
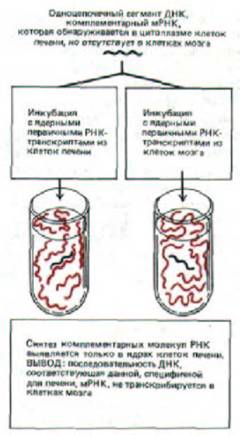
Рис. 10-5. Схема эксперимента, демонстрирующего, что экспрессия генов в клетках млекопитающих контролируется главным образом на уровне транскрипции (см. рис. 10-4).
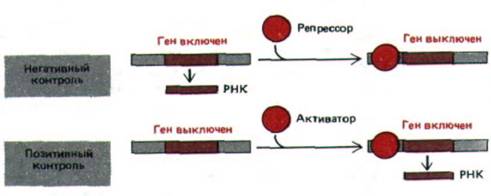
Рис. 10-6. Сравнение позитивной и негативной регуляции. В данном примере показан контроль на уровне транскрипции, однако те же два типа контроля могут быть задействованы на любом из этапов регуляции, представленных на рис. 10-2.
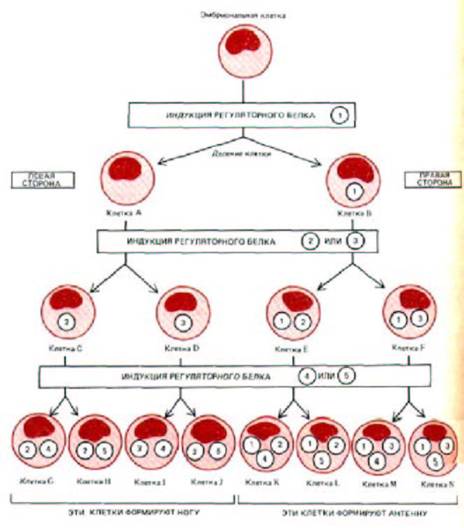
Рис. 10-7. Упрощенная схема дифференцировки клеток эмбриона, иллюстрирующая, каким образом разные комбинации небольшого числа белков-регуляторов могут обеспечить возникновение клеток многих типов.
Эта предельно упрощенная схема предполагает, что после каждого деления происходит выбор в пользу синтеза какого-либо одного из двух белков-регуляторов (обозначенных пронумерованными кружочками). При этом сообразно занимаемой условной позиции дочерние клетки, отходящие влево относительно общего плана строения эмбриона (и относительно страницы этой книги), всегда индуцируют синтез белков с четными номерами, тогда как дочерние клетки, отходящие вправо-синтез белков с нечетными номерами. Считается, что синтез каждого белка-
регулятора - самоподдерживающийся процесс (см. рис. 10-33 и 10-35). Таким образом, клетки разрастающегося клона будут содержать все увеличивающееся число различных белков-регуляторов, каждый из которых контролирует целую «батарею» генов.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
зародыша дрозофилы. Набор регуляторных генов определяет последовательное подразделение эмбриона на отличающиеся друг от друга участки (см. разд. 16.5.5).
10.1.6. Активность гена обычно зависит от действия нескольких регуляторных белков [5]
На первый взгляд схема комбинационной регуляции активности генов, представленная на рис. 10-7, дает основание для вывода о постепенно накапливающихся различиях между клетками последующих поколений. Например, можно предположить, что добавление регуляторного белка 2 к клеткам С и Е приведет к появлению в этих клетках одного и того же набора дополнительных белков (тех, которые кодируются генами, активируемыми белком-регулятором 2). Подобная точка зрения неверна по очень простой причине. Комбинационная регуляция гена гораздо сложнее этой схемы потому, что различные регуляторные белки взаимодействуют друг с другом. Даже у бактерий для включения одного-единственного гена иногда бывает необходимо взаимодействие двух различных регуляторных белков (см. разд. 10.2.2). У высших эукариот транскрипция какого-либо гена обычно требует совместного действия целого кластера активаторных белков (см. разд. 10.2.9).
Например, белок 2 при взаимодействии с активаторным белком 1 может включать в клетке Е иной набор генов, нежели тот, который он включает в клетке С. По-видимому, именно поэтому единственный белок-рецептор стероидного гормона (пример белка-регулятора) в различных типах клеток млекопитающих определяет синтез разных наборов белка (см. разд. 12.2.2). В целом, специфические изменения в экспрессии гена, возникающие в результате синтеза регуляторного белка, зависят от предыстории клеток, так как именно эти предыдущие события и определяют, какие белки-
регуляторы уже имеются в клетке (рис. 10-8).
10.1.7. Главные белки-регуляторы активируют сразу много генов [6]
Как отмечалось ранее, в клетке содержится много регуляторных белков, каждый из которых, действуя в комбинации с другими белками этого класса, контролирует многочисленные гены. Вероятно, существует разветвленная схема взаимодействий белков, согласно которой каждый
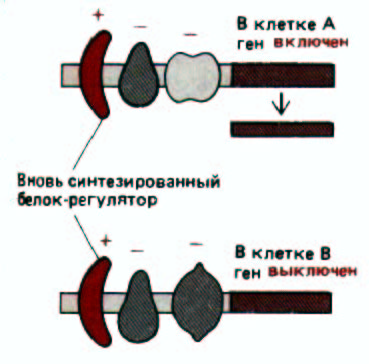
Рис. 10-8. Воздействие вновь синтезированных белков-регуляторов на клетку. Подобное воздействие зависит от того, какие белки-
регуляторы уже имеются в клетке и, следовательно, от ее предыстории. На схеме один и тот же ген изображен в клетках А и В. Изначально этот ген в обеих клетках выключен. Однако в клетке А продуцируется белок, изображенный крайним слева и отсутствующий в клетке В. Для простоты принимается, что каждый белок-регулятор либо положительно, либо отрицательно воздействует на транскрипцию, которая определяется их совместным воздействием. В действительности же общее воздействие не обязательно осуществляется по принципу аддитивности. Например, в некоторых случаях два белка-регулятора при связывании с ДНК взаимодействуют друг с другом, причем происходит изменение активности каждого из них.
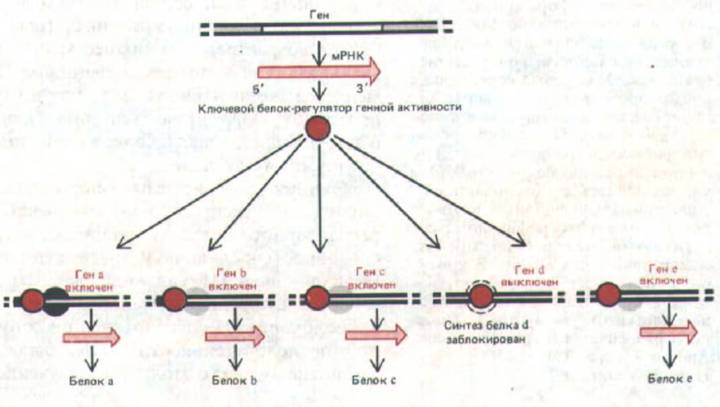
Рис. 10-9. Схема, иллюстрирующая, как «решение» синтезировать один-единственный главный белок-регулятор может воздействовать на образование самых разных белков в клетке.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
из регуляторных белков контролирует гены, кодирующие другие белки-регуляторы и так далее.
Однако не все регуляторные белки равны. Некоторые из них (главные белки-регуляторы) обладают решающим, координирующим действием на активность многих генов (рис. 10-9). Например, в гл. 16 будет показано, как ряд мутаций одного-единственного гена у Drosophila превращает одну часть тела мухи в другую. Мутации, вызывающие подобные изменения, известны под названием гомеотических (или гомеозисных). Одна из таких мутаций, названная Antennapedia, обусловливает синтез абберантного главного белка-регулятора. В результате целая группа клеток, в норме образующих антенну, коренным образом меняет свое поведение и формирует ногу; в итоге на свет появляется муха с растущей на голове ногой. О существовании главных регуляторных белков у позвоночных (включая человека) может свидетельствовать следующий факт. Отсутствие одного-единственного белка (рецептора стероидного гормона тестостерона) приводит к тому, что эмбрион с мужским генотипом (XY) развивается в фенотипически почти нормальную женщину (см. разд. 12.2.3). Прямое доказательство участия главных регуляторных белков в развитии позвоночных получено недавно при исследовании клеток скелетной мышцы.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 698; Нарушение авторских прав?; Мы поможем в написании вашей работы!