КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Технологія рекомбінантних ДНК
|
|
|
|
Трансляція
Метоболізму
Транскрипція
У генах закодована інформація про білки, що синтезуються в клітині. Однак, сама ДНК не використовується в якості безпосередньої матриці для синтезу білка. Реалізація генетичної інформації становить двостадійний процес. На першій стадії ген є матрицею для синтезу молекул РНК, на які досконало транскрибується (переписується) послідовність нуклеотидів відповідного гена і, отже, інформація про послідовність амінокислот, що закодована в ньому. На другій стадії нуклеотидна послідовність РНК транслюється (перекладається) у поліпептидний ланцюг.
Таким чином, потік генетичної інформації у клітині здійснюється за таким напрямом:
ТранскрипціяТрансляція
ДНК РНК білок регуляція
РНК − це лінійна полінуклеотидна молекула, що відрізняється від ДНК за двома відношеннями. По-перше, моноцукром у РНК є рибоза, що містить не одну, а дві гідроксильні групи; вони пов'язані з 2'- і З'-атомами вуглецю. По-друге, одним із чотирьох основ у РНК є урацил (U), що займає місце тиміну. Більшість молекул РНК одноланцюгова, хоча часто в них є взаємкомплементарні ділянки, що утворюють дволанцюгові структури − «шпильки». Спарювання основ відбувається таким чином, як і в ДНК, за винятком того, що замість пари А-Т утворюється A-U.
Існують три основних типи РНК: інформаційна, або матрична (ІРНК, мРНК), яка є матрицею для синтезу білка. Кожному структурному гену (або групі генів) відповідає власна молекула мРНК; транспортна (тРНК), що переносить амінокислоти в активованій формі до рибосом, де здійснюється синтез молекули білка; рибосомна (рРНК) − необхідний компонент рибосом. Усі вони відіграють важливу роль у процесі розшифровки генетичної інформації. У більшості прокаріотів транскрипція всіх РНК здійснюється за допомогою одного ферменту − РНК-полімерази. В еукаріот мРНК, рРНК і тРНК транскрибуються різними РНК-полімеразами.
Транскрипція багато в чому подібна до реплікації. Матрицею при синтезі РНК є певна ділянка одного з ланцюгів ДНК. РНК-полімераза копіює цю ділянку, послідовно з'єднуючи рибонуклеотиди один з одним за допомогою 3'-5'-фосфодіефірних зв'язків відповідно до правила комплементарності. Транскрипція починається після приєднання РНК-полімерази до специфічної нуклеотидної послідовності − промотору. Завершується транскрипція коли РНК-полімераза досягає послідовності стоп-сигналу, або сигналу термінації транскрипції. Ділянка ДНК, що обмежена промотором і стоп-сигналом, становить одиницю транскрипції − транскриптон. Дроблення ДНК на множину транскриптонів забезпечує можливість незалежного зчитування різних генів, їх індивідуального вмикання і вимикання.
У ході транскрипції новосинтезована молекула РНК від'єднується від ДНК і подвійна спіраль ДНК відновлюється. Щоб забезпечити транскрипцію тільки окремих сегментів ДНК, повинні існувати якісь сигнальні послідовності, що вказують, де починається (ініціюється) транcкрипція й де вона зупиняється (термінується). Сигнал ініціації звичайно розташовується перед послідовністю, що кодує, а сигнал термінації − слідом за нею. Ділянка ДНК, що передує гену, який транскрибується, називається 5'-фланкуючою послідовністю, а розташована за ним − 3'-фланкуючою.
Цикл транскрипції можна поділити на чотири основні стадії, до кожної з яких входить багато елементарних подій:
· зв'язування з ДНК;
· ініціація ланцюга РНК;
· ріст (елонгація) ланцюга РНК;
· термінація (припинення) росту ланцюга РНК.
Цикл транскрипції починається із приєднання РНК-полімерази до промотору − певної ділянки ДНК, що визначає місце початку синтезу РНК. При зв'язуванні РНК-полімерази з промотором створюється так званий закритий промоторний комплекс, в якому ДНК зберігає двоспіральну структуру. Закритий комплекс може обертатися у відкритий, причому цей процес − зворотний. Після утворення відкритого комплексу РНК-полімераза розплітає приблизно один виток подвійної спіралі ДНК в районі стартової точки − нуклеотиду, з якого починається комплементарне копіювання матриці. У відкритому комплексі зв'язок РНК-полімерази із ДНК суттєво зміцнюється.
Наступна стадія, ініціація, здійснюється за наявністю субстратів РНК-полімерази, нуклеозидтрифосфатів і полягає в утворенні перших декількох ланок ланцюга РНК. Ріст ланцюга РНК відбувається в напрямку 5'  3', як і під час реплікації ДНК, а РНК-полімераза пересувається вздовж матричного ланцюга в напрямку 3' 5'.
3', як і під час реплікації ДНК, а РНК-полімераза пересувається вздовж матричного ланцюга в напрямку 3' 5'.
Ефективність ініціації на різних промоторах, їх «сила», суттєво розрізняється: з деяких промоторів за період поділу клітини може ініціюватися всього одна-дві молекули РНК, в той час як на інших −ініціювання відбувається кожні одну-дві секунди.
На стадії елонгації в ДНК розплетено біля 18 п.н. Близько 12 нуклеотидів матричної нитки ДНК створює гібридну спіраль з наростаючим кінцем ланцюга РНК. Відповідно до руху РНК-полімерази вздовж матриці попереду неї здійснюється розплітання, апозаду − відновлення подвійної спіралі ДНК. Одночасно відбувається звільнення синтезованої РНК із комплексу з матрицею і з РНК-полімеразою. Максимальна швидкість елонгації становить близько 50 нуклеотидів за секунду.
На відміну від ДНК-полімерази РНК-полімераза не володіє здатністю до самокорекції. У зв'язку з цим надійність транскрипції значно нижча, ніж надійність реплікації. Частота помилок під час синтезу РНК дорівнює: одна на 106 нуклеотидів. Низька надійність синтезу РНК компенсується клітиною значною кількістю копій РНК-транскриптів з одного гена.
Термінація транскрипції також ретельно регулюється, як й ініціація. За відсутності спеціальних білкових факторів термінації РНК-полімераза здатна зупиняти синтез РНК на тих термінаторах, нуклеотидна послідовність в районі яких відрізняється двома особливостями. По-перше, за ходом транскрипції знаходиться ділянка, яка містить багато GCнуклеотидів і для неї здатна центральна симетрія, по-друге, слідом йде ділянка, що складається з 4... 8 розташованих поряд аденінів (полі-А). Транскрипція закінчується наприкінці полі-А послідовності, або одразу після неї. Вважають, що після проходження РНК-полімеразою першої ділянки, в РНК-продуктів виникає «шпилька», що призводить до зупинки ферменту і звільнення РНК-продукту.
Регуляція транскрипції. Всі процеси, що відбуваються в бактеріальній клітині, − створення амінокислот, нуклеотидів, реплікація, трансляція, звільнення енергії − потребують участі білків. Однак, енергетичних ресурсів клітини не вистачає для одночасного здійснення транскрипції і трансляції всіх структурних генів (гени, в яких закодована інформація про структуру кінцевого продукту). Тому постійно відбувається експресія лише тих генів, які кодують білки, що підтримують основні клітинні функції, а транскрипція інших структурних генів регулюється.
Регуляція транскрипції у бактерій. У бактерій часто білки одного метаболічного шляху кодуються суміжними структурними генами. Нуклеотидна послідовність, у якої закодовано більше одного білка, називається опероном. Звичайно оперон перебуває під контролем єдиного промотору, і при його транскрипції утворюється одна довга молекула мРНК, що кодує кілька білків. За трансляції такої мРНК, у якої стоп-кодон послідовності, що кодує один білок, є сосудом зі старт-кодоном гена наступного білка, синтезується набір білків.
У більшості структурних генів Е. соli є два сайти зв'язку для РНК-полімерази. Один з них, звичайно, являє собою нуклеотидну послідовність
ТАТААТ
АТАТТА
(ТАТА-бокс, або бокс Прибнова), а інший −
TTGAC
AACTG.
ТАТА-бокс і послідовність TTGACрозташовані за 10 (ділянка -10) і 35 (ділянка -35) нуклеотидів до сайту ініціації транскрипції відповідно (нуклеотид +1). Звичайно, від ділянки між ТАТА-боксом і нуклеотидом +1 набагато залежить, чи буде відбуватися транскрипція даного оперона. Для вмикання й вимикання різних оперонів у ході еволюції сформувалася безліч регуляторних систем.
Відповідно до моделі оперона нуклеотидна послідовність містить, якнайменш, чотири компоненти регулювання: структурний ген (або гени, що контролюють взаємозв'язані біохімічні функції S1, S2, S3), ген-регулятор (R), ген-оператор (О), ген-промотор (Р) (рис. 8).

Рис. 8. Модель оперона:
R− ген регулятор, Р − ген промотор,О − ген оператор, S1, S2, S3- структурні гени, t– термінуючаділянка,Y− речовина-індуктор
Ген-регулятор визначає структуру білка репресора. Білок-репресор здатний зв'язуватися з геном-оператором (О), у цьому разі процес зчитування інформації припиняється, оскільки РНК-полімераза не може пересуватися вздовж молекули ДНК. Ген-оператор (О) відповідає за порядок зчитування інформації, ген-промотор (Р) є початковою ділянкою для зв'язування РНК-полімерази, ферменту, який каталізує транскрипцію ДНК в мРНК.
Якщо у середовище додати речовину-індуктор (Y), то білок репресор блокується, втрачає здатність з'єднуватися з геном-оператором і процес транскрипції здійснюється. Звичайно індуктор руйнується клітинними ферментами. Коли його концентрація знижується, репресор зв'язується з операторною ділянкою, і транскрипція знову припиняється. Операторна ділянка специфічна для кожного оперона, а індуктор взаємодіє тільки з певним репресором.
У разі, коли необхідно, щоб експресія відбувалася постійно, проводять мутації в гені-регуляторі, тоді змінюється структура білка-репресора і він втрачає здатність зв'язуватися з геном-оператором, або мутацію здійснюють у гені-операторі, тоді білок-репресор не розпізнає його і зв'язок також не відбувається. Такі мутанти мають назву конститутивні мутанти.
Регуляція транскрипції у еукаріот. У еукаріот більшість структурних генів складається з декількох дискретних ділянок, що кодують (екзонів), розділених ділянками, що не кодують (інтронами). По завершенні транскрипції еукаріотичного структурного гена інтрони вирізають із первинного продукту транскрипції за допомогою ферментів (процесинг), а екзони скріплюються один з одним «торець у торець» (сплайсшг) з утворенням функціональної мРНК (рис. 9). Звичайно довжина екзонів становить від 150 до 200 нуклеотидів, а довжина інтронів варіює від 40 до 10000 нуклеотидів. Деякі еукаріотичні структурні гени взагалі не мають інтронів.
Іноді сплайсинг мРНК може проходити альтернативним шляхом. Наприклад, в одній тканині функціональна мРНК може утворюватися за з'єднання всіх екзонів первинного транскрипта, а в іншій якийсь екзон буде вирізаний разом із фланкуючими його інтронами й утвориться інша функціональна мРНК. Завдяки альтернативному сплайсингу в різних тканинах можуть утворюватися різні продукти того самого структурного гена (рис. 10,11).
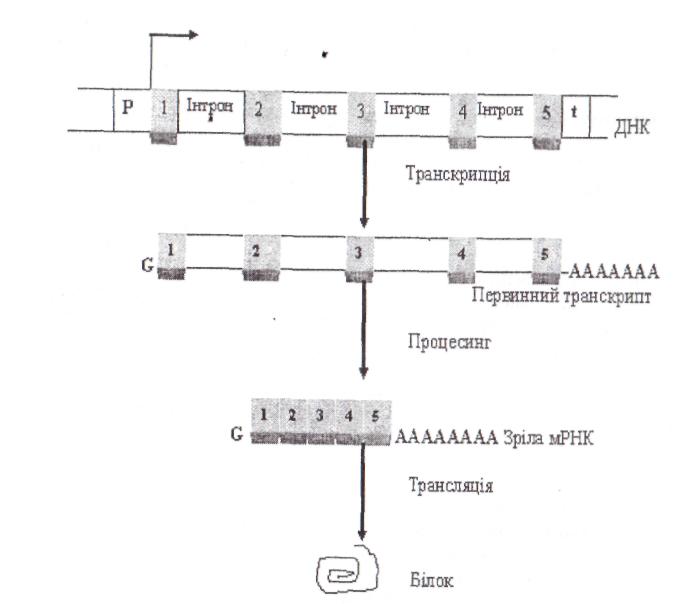
Рис. 9 . Схематичний процес реалізації інформації еукаріотичного структурного гена: 1, 2, 3, 4, 5 − структурні гени
Для включення й вимикання транскрипції різних еукаріотичних структурних генів використовується безліч різноманітних високоспецифічних процесів. Але так чи інакше регуляція транскрипції в еукаріот здійснюється за допомогою специфічних білків і факторів транскрипції. Багато хто з них зв'язується безпосередньо з нуклеотидною послідовністю завдовжки менше 10 п.н., що називається регуляторним елементом. На відміну від прокаріот в еукаріот оперони здебільшого відсутні, тобто кожний еукаріотичний структурний ген має власний набір регуляторних елементів. Істотну роль у регуляції транскрипції в еукаріот, крім опосередкованої взаємодією між ДНК і білками, грають також білок −білкові взаємодії.

Рис. 11. Альтернативний сплайсинг первинного транскрипту в еукаріот
Незважаючи на індивідуальність набору регуляторних елементів у структурних генів еукаріот, кожен з них має промоторну ділянку (ТАТА-бокс, або бокс Хогнесса) з восьми нуклеотидів, що включає послідовність ТАТА; послідовність ССААТ (САТ-бокс); ділянку з повторюваних дінуклеотидів GC (GC-бокс). Ці елементи перебувають на відстані 25, 75 і 90 п.н. від сайту ініціації, відповідно. Транскрипція структурного гена еукаріот починається зі зв'язування із ТАТА-боксом фактора транскрипції, що являє собою комплекс, принаймні, з 14 білків. Надалі з ними зв'язуються інші фактори транскрипції, і, нарешті, з усім цим транскрипційним комплексом зв'язується РНК-полімераза II. Далі, за участі додаткових факторів, відбувається ініціація транскрипції.
Трансляція − це процес декодування мРНК, внаслідок якого інформація з мови послідовності основ мРНК переводиться на мову амінокислотної послідовності білка. В цьому процесі погоджено взаємодіють більше сотні видів макромолекул. Крім рибосом необхідні молекули тРНК, ферменти, що активують, розчинні фактори та мРНК.
У клітині, що активно синтезує білки, містяться до 60 різних видів тРНК. тРНК − це лінійна одноланцюгова молекула завдовжки від 75 до 93 нуклеотидів. У ній є декілька взаємокомплементарних ділянок, що спаровуються між собою. За допомогою специфічних ферментів (аміноацил-тРНК − синтетаз) до 3'-кінця тРНК приєднується відповідна амінокислота. Для кожної із двадцяти амінокислот, з яких складаються всі білки, існує принаймні одна специфічна тРНК. На іншому кінці молекули тРНК розташована послідовність із трьох нуклеотидів, що називається антикодоном. Вона розпізнає специфічний кодон у мРНК і визначає, яка саме амінокислота буде приєднана до зростаючого поліпептидного ланцюга (рис. 12).
Синтез білка здійснюється шляхом послідовної поліконденсації окремих амінокислотних залишків, починаючи з аміно-(Ь0-кінця поліпептидного ланцюга у напрямку карбоксильного (С)-кінця. Транскрипції передує активація тРНК − приєднання амінокислоти до З'-кінцевого аденозину молекули тРНК з утворенням аміноацил-тРНК.

Рис. 12. Будова молекули тРНК
Синтез білка відбувається за три стадії:
1. −ініціація забезпечує зв'язування ініціаторної тРНК із сигналом початку трансляції в мРНК, при цьому ініціаторна тРНК займає Р-ділянку рибосоми, одну з двох ділянок зв'язування тРНК;
2. − елонгація починається зі зв'язування аміноацил-тРНК з іншою ділянкою зв'язку на рибосомі (А-ділянкою) з наступним утворенням пептидного зв'язку із другої аміноацил-тРНК, що поєднана з ініціаторною тРНК. Діпептидил-тРНК пересувається з А-ділянки у Р-ділянку, нова амінокислота зв'язується з А- ділянкою, що звільнилася, і починається новий цикл реакціїелонгації;
3. − термінація відбувається коли стоп-кодон (сигнал термінації) вмолекулі тРНК зчитується фактором звільнення білка, щовикликає відокремлення завершеного поліпептидного ланцюгавід рибосоми.
Яким чином молекула тРНК розпізнає власну амінокислоту? Існує спеціальний набір ферментів, так званих аміноацил-тРНК-синтетаз, що приєднують амінокислоти до відповідних молекул тРНК. Кожній з амінокислот відповідає власна особлива синтетаза: вона приєднує аденін до аденінової тРНК, гліцин − до гліцинової та ін.
Аміноацил-тРНК-синтетаза виконує роль адаптера, здійснює високоспецифічну підгонку однієї молекулярної поверхні до іншої.
Молекули тРНК відіграють роль кінцевих адаптерів за переведення інформації, що закодована в нуклеотидній послідовності, на мову білка. Таким чином, генетичний код розшифровується за допомогою двох взаємопов'язаних наборів адаптерів.
Матрична мРНК може бути представлена десятками різних типів молекул, а рРНК − усього двома типами. Більша рРНК утворює із білками рибонуклеопротеїдний комплекс, названий великою рибосомною субодиницею, а рРНК меншого розміру комплекс, названий малою рибосомною субодиницею. Під час синтезу білків субодиниці поєднуються з утворенням рибосоми. В еукаріот обидві рибосомні субодиниці крупніші, ніж у прокаріот.
Усі етапи білкового синтезу стають можливими внаслідок того, що трансляція здійснюється великим мультиферментним комплексом − рибосомою, що складається з молекул білків і РНК. Протягом усього процесу синтезу білка зростаючий поліпептидний ланцюг, мРНК і чергова аміноацил-тРНК залишаються прикріпленими до рибосоми. В рибосомах обох типів є борозенка, яка утримує зростаючий поліпептидний ланцюг, і борозенка, що утримує молекулу мРНК. Довжина борозенок така, що у першої розміщується до 30 амінокислот, а у другої − до 35 нуклеотидів РНК.
У рибосомі є дві ділянки, що зв'язують молекули тРНК. Одна з них утримує молекулу тРНК, що приєднана до зростаючого кінця поліпептидного ланцюга; тому її називають пептидил-тРНК-сполучною ділянкою, або Р-ділянкою. Інша затримує щойно прибулумолекулу тРНК, що навантажена амінокислотою; її називають аміноацил-тРНК-сполучною ділянкою, або А-ділянкою. До обох ділянок молекула тРНК прикріпляється лише в тому разі, якщо її антикодон парується із комплементарним до нього кодоном мРНК. А-і Р-ділянки розташовані дуже близько одна до одної і так, що дві пов'язані з ними молекули тРНК спаровуються із двома сусідніми кодонами в молекулі мРНК
Якщо А-ділянка рибосоми занята одним із стоп-кодонів: UAA, UGA або UAG, то спаровування аміноацил-тРНК, як правило, не відбувається. Нормальні клітини не містять тРНК з антикодонами, комплементарними сигналам термінації. Останні розпізнаються білковими факторами звільнення, що свідчить про те, що білки здатні розпізнавати тринуклеотидні послідовності з високим ступенем точності.
Зв'язування фактора звільнення змінює активність пептидилтрансферази, тому вона тепер приєднує до пептидил-тРНК не вільну аміногрупу амінокислоти, а молекулу води. Внаслідок цього карбоксильний кінець поліпептидного ланцюга відокремлюється від молекули тРНК. Оскільки зростаючий поліпептид утримується на рибосомі лише за рахунок зв'язку з молекулою тРНК, завершений білковий ланцюг виявляється звільненим і, після відокремлення від рибосоми, потрапляє у цитоплазму.
Технологія рекомбінантних ДНК (її називають такожмолекулярним клонуванням або генною інженерією) − це сукупністьекспериментальних процедур, що дозволяє здійснювати перенесення генетичного матеріалу (ДНК) з одного організму до іншого. Ніякогоєдиного, універсального набору методик тут не існує, але найчастішеексперименти з рекомбінантною ДНК здійснюють за такою схемою:
−з організму − донора потрібних генів - екстрагують чужоріднуДНК, піддають її ферментативному гідролізу (розщеплюють,розрізають) із'єднують (лігують, скріплюють) з іншою ДНК(вектора для клонування) з утворенням нової, рекомбінантноїмолекули;
−цю конструкцію вводять у клітину-реципієнт, де відбувається її реплікація й передача нащадкам. Цей процес називається трансформацією;
−ідентифікують і відбирають клітини, що несуть рекомбінантну ДНК (трансформовані клітини);
−одержують специфічний білковий продукт, синтезований клітинами-реципієнтами, що є підтвердженням клонування шуканого гена.
Необхідність маніпулювання генами диктується конкретними завданнями фундаментальних і прикладних досліджень. Для розуміння молекулярних механізмів функціонування окремих генів і взаємозалежних генетичних систем велике значення має робота з ізольованими генами. Такі дослідження дозволяють визначити границі генів, виділити їх у чистому вигляді й ідентифікувати елементи структури, істотні для функціонування. Доказом функціональної значимості виділеної ділянки генома може бути тільки його нормальна експресія в модельній генетичній системі. Тому наступним етапом дослідження виділеного гена завжди є переміщення його в таку генетичну систему, де експресія гена легко виявляється. Результати експресії оцінюють або за появою білкового продукту, що кодується досліджуваним геном, або за зміною функцій біологічної системи внаслідок появи в ній нової ферментативної або іншої активності.
У цей час за допомогою методів генної інженерії отримано дані про структуру й функціонування генів різноманітних організмів, що дало можливість перейти на якісно новий рівень генетичних досліджень. Це, по-перше, можливість перенесення гена в нове для нього генетичне оточення з подальшою його експресією, що веде до зміни властивостей організму, у геном якого вводиться ген (наприклад, створення продуцентів біологічно активних речовин або трансгенних тварин), а також здійснення генотерапії спадкових і придбаних захворювань шляхом штучного заміщення мутаційних алелей. По-друге, стало реальним конструювання нових генів шляхом об'єднання invitro як відомих, так і нових, штучно синтезованих послідовностей нуклеотидів. Цей підхід використовується в білковій інженерії для дослідження функціональної значимості окремих амінокислот і доменів у поліпептидних ланцюгах ферментів, а також для створення нових білків. По-третє, у сучасній біотехнології з'явилася можливість застосовувати ізольовані гени в складі генно-інженерних конструкцій для одержання продуктів харчування і біологічно активних речовин білкової природи.
Оскільки в експериментальних умовах неможливо працювати з однією копією гена, одержання необхідного числа ідентичних копій гена або його частин є першим і одним із основних завдань генної інженерії. Для його вирішення використовують метод молекулярного клонування. Сутність методу полягає в тому, що нуклеотидна послідовність, яку необхідно виділити або розмножити, ковалентно вбудовується в молекули нуклеїнової кислоти, які здатні до самореплікації, і називаються векторами. Далі така послідовність нуклеотидів у складі вектора вводиться в клітини про- або еукаріотичного організму, і ці гібридні клітини в селективних умовах, що забезпечують збереження вектора усередині клітин, вирощують на поживному середовищі. У результаті створюється клон клітин, теоретично утримуючих ідентичні векторні молекули з однією й тією ж вставкою чужорідної послідовності нуклеотидів. Оскільки об'єднання молекул послідовності нуклеотидів, що клонуються, і вектора є не чим іншим, як рекомбінацією invitro, такі гібридні молекули називають рекомбінантними молекулами. У наш час розроблено численні методи, що дозволяють виділяти певні послідовності нуклеотидів зі складної суміші фрагментів хромосомної ДНК, а також здійснювати обмін між точно визначеними фрагментами генів й інших послідовностей нуклеїнових кислот. У всіх цих реакціях, як правило, використовуються високоочищені препарати нуклеїнових кислот і ферментів нуклеїнового обміну.
Конструювання рекомбінантних молекул здійснюється за допомогою цілого арсеналу ферментів − обов'язкового й незамінного інструменту практично всіх етапів цього складного процесу.
|
|
|
|
|
Дата добавления: 2017-02-01; Просмотров: 212; Нарушение авторских прав?; Мы поможем в написании вашей работы!