КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Биосинтез углеводов
|
|
|
|
Некоторые общие организационные принципы биосинтеза:
1. Пути биосинтеза и пути расщепления тех или иных биомолекул, как правило, не идентичны. Эти пути могут включать общие обратимые реакции, но у них всегда имеется хотя бы одна ферментативная реакция, по которой они различаются.
2. Биосинтетические пути и соответствующие им катаболические пути контролируются разными регуляторными ферментами.
3. Требующие затраты энергии биосинтетические процессы обязательно сопряжены с поставляющим энергию расщеплением АТФ, вследствие чего весь процесс в целом является практически необратимым, точно так же как в целом необратим катаболизм.
У всех высших животных биосинтез D-глюкозы – абсолютно необходимый процесс, потому что D-глюкоза крови служит единственным или главным источником топлива для нервной системы, почек, семенников, эритроцитов и для всех клеток эмбриона. У человека один только мозг потребляет более 120 г глюкозы в сутки.
Гликоген печени служит резервуаром глюкозы: из него образуется глюкоза, которая поступает в кровь. Мышечный гликоген служит источником энергии для мышечного сокращения. У животных образование глюкозы из неуглеводных предшественников называется глюконеогенез. Важные предшественники: лактат, пируват, глицерол, большинство аминокислот и промежуточные продукты цикла Кребса. У животных глюконеогенез протекает главным образом в печени и значительно меньше – к корковом слое почек.
Центральный путь глюконеогенеза – превращение пирувата в глюкозу. Семь легко обратимых реакций гликолиза свойственны также и глюконеогенезу. Три практически необратимых реакций гликолиза имеют обходные пути в глюконеогенезе.
Первый обходной путь – превращение пирувата в фосфоенолпируват, протекает в несколько реакций. Первую реакцию катализирует пируваткарбоксилаза – биотинсодержащий фермент митохондрий, ее активатором служит ацетил-СоА, в его отсутствие фермент полностью лишен активности:
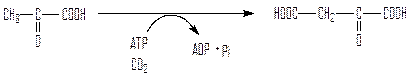
пируват оксалоацетат
Вторая реакция катализируется малатдегидрогеназой цикла Кребса:
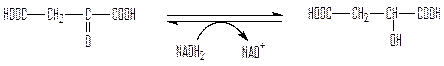
оксалоацетат малат
Малат покидает митохондрии при участии специальной дикарбоксилатной транспортной системы, находящейся во внутренней митохондриальной мембране, и поступает в цитозоль. Здесь он окисляется под действием цитозольной формы НАД-зависимой малатдегидрогеназы с образованием внемитохондриального оксалоацетата:
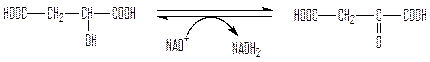
малат оксалоацетат
Далее фосфоенолпируваткарбоксикиназа (не путать с фосфоенолпируваткарбоксилазой С4-растений!) фосфорилирует оксалоацетат за счет энергии ГТФ с одновременным отщеплением СО2:
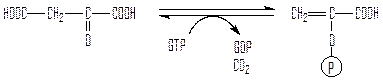
оксалоацетат фосфоенолпируват
На фосфорилирование одной молекулы пирувата до фосфоенолпирувата затрачивается энергия двух высокоэнергетических фосфатных групп. Когда фосфоенолпируват превращается в пируват в процессе гликолиза, из АДФ синтезируется только одна молекула АТФ.
Второй обходной путь – превращение фруктозо-1,6-дифосфата во фруктозо-6-фосфат под действием фруктозодифосфатазы:
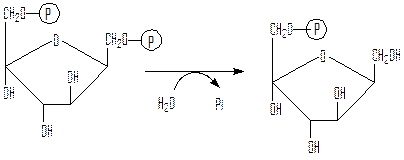
фруктозо-1,6-дифосфат фруктозо-6-фосфат
Фруктозодифосфатаза – регуляторный фермент, ингибитором для нее выступает АМФ, а активатором – АТФ.
Третий обходной путь – превращение глюкозо-6-фосфата в свободную глюкозу под действием фермента глюкозо-6-фосфатазы:
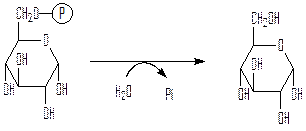
глюкозо-6-фосфат глюкоза
Глюкозо-6-фосфатаза обнаруживается только в той фракции клеток печени позвоночных, которая содержит эндоплазматический ретикулум. Глюкозо-6-фосфатаза отсутствует в таких тканях, как мышцы или мозг, так что они не поставляют в кровь свободную глюкозу.
Синтез глюкозы из пирувата обходится организму довольно дорого. Однако немалая часть этой платы расходуется лишь на то, чтобы обеспечить необратимость глюконеогенеза. Суммарная реакция глюконеогенеза:
2Пируват + 4АТФ + 2ГТФ + 2НАДН2 + 4Н2О → Глюкоза + 4АДФ + 2ГДФ + 6Pi + 2НАД+.
Регуляция глюконеогенеза:
А) Биосинтез глюкозы из пирувата усиливается когда в клетке накапливается больше митохондриального ацетил-СоА, чем ей нужно для цикла Кребса. Накопившийся ацетил-СоА ингибирует пируватдегидрогеназный комплекс и способствует биосинтезу глюкозы из пирувата.
Б) Когда клетка полностью обеспечена АТФ, то активируется фермент фруктозодифосфатаза и ингибируется фосфофруктокиназа..
В) Гормональная регуляция.
Г) Непрямая регуляция – через пируваткиназу гликолиза. Пируваткиназа существует в двух формах – L (liver – печень) и М (muscle – мышца). L-форма преобладает в тканях, способных к глюконеогенезу, и аллостерически ингибируется избытком АТФ и некоторыми аминокислотами (аланином). М-пируваткиназа такой регуляции не подчиняется.
Ацетил-СоА никогда в животных клетках не превращается в пируват, поэтому жирные кислоты с четным количеством атомов углерода не превращаются в глюкозу.
Путь биосинтеза гликогена отличается от пути его расщепления. Сначала гексокиназа фосфорилирует за счет АТФ глюкозу с превращением ее в глюкозо-6-фосфат (первая реакция гликолиза). Далее фосфоглюкомутаза превращает глюкозо-6-фосфат в глюкозо-1-фосфат (реакция из гликогенолиза). Далее идет ключевая реакция биосинтеза гликогена, отсутствующая в процессе его расщепления:
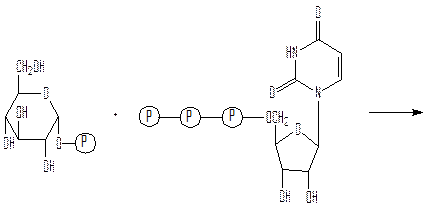
глюкозо-1-фосфат УТФ
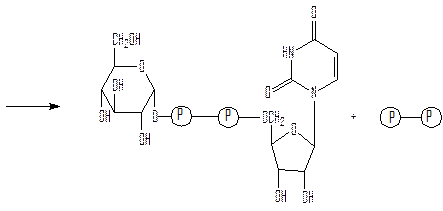
уридиндифосфатглюкоза (УДФ-глюкоза) пирофосфат
Реакция вынуждена идти слева направо под действием пирофосфатазы, гидролизующей пирофосфат (PPi) до фосфата (Pi). УДФ-глюкоза играет роль промежуточного продукта в процессе образования гликогена. Перенос гликозильных групп от УДФ-глюкозы на нередуцирующий конец разветвленной молекулы гликогена катализируется гликогенсинтазой. В этой реакции образуется новая α(1→4) связь. Гликогенсинтазе требуется в качестве затравки α(1→4) полиглюкозная цепь, состоящая не менее чем из четырех глюкозных остатков, к которым фермент последовательно присоединяет гликозильные группы с нередуцирующего конца.

Гликогенсинтаза неспособна катализировать образование α(1→6) связей, находящихся в точках ветвления цепей гликогена. Существует специальный «ветвящий» фермент – гликозил-(4→6)-трансфераза. Он катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6-7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксильную группу остатка глюкозы той же или другой цепи гликогена, расположенного ближе к внутренней части молекулы. В результате этого образуется новая боковая цепь. После этого гликогенсинтаза может добавлять к этой боковой цепи новые остатки глюкозы.
Биологический смысл ветвления заключается в повышении растворимости гликогена и в увеличении числа нередуцирующих концов у его молекулы, что делает гликоген более доступным для действия гликогенфосфорилазы и гликогенсинтазы.
Крахмал в растениях синтезируется таким же путем, но донором гликозильных групп служит при этом не УДФ-глюкоза, а АДФ-глюкоза.
Гликогенсинтаза и гликогенфосфорилаза регулируются реципрокно, то есть прямо противоположным образом.
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 1515; Нарушение авторских прав?; Мы поможем в написании вашей работы!