КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физиология дыхания рыб 1 страница
|
|
|
|
Р-деполяризация предсердий,Q- начало деполяризации желудочка,R-основной зубец деполяризации желудочка, S- окончание деполяризации,QRS-совакупность зубцов отражающих деполяризацию желудочка,T-реполяризация желудочка,P-Q- плато ПД предсердии. Начало деполяризации проводящей системы желудочка (предсердный комплекс), Q T – желудочковый комплекс, T P-пауза
1 2 3
Рис. 26. Фазы сокращения сердца: 1- общая пауза;2-сокращение предсердия; 3 - сокращение желудочка
Ритм сердца или ритмическая деятельность сердца -образует повторяющиеся циклы - совокупность электрических, механических и биохимических процессов в течение одного полного сокращения и расслабления.
Последовательность сокращений сердца у позвоночных по существу одинакова, независимо от числа его камер. Во время сокращения желудочка, когда аортальные (полулунные) клапаны открыты, аорта наполняется кровью, а сердце опорожняется. Систола желудочка заканчивается, когда давление в желудочке падает ниже уровня давления в аорте, и аортальные клапаны закрываются. Во время сокращения желудочка давление в предсердии ниже, чем в желудочке, а атриовентринулярные клапаны остаются закрытыми.
Сердце рыб гораздо меньше и слабее, чем сердце наземных позвоночных. Масса его обычно не превышает 2,5%, в среднем 1% массы тела, тогда как у млекопитающих оно достигает 4,6%, а у птиц даже 16%. Поэтому сердце должно обеспечить давление, достаточное для проталкивания крови последовательно через две капиллярные сети. Кровяное давление (Па) у рыб низкое - 2133,1 (скат), 11198,8 (щука), 15998,4 (лосось), тогда как в сонной артерии лошади - 20664,6.
Определение частоты сокращений сердца, вероятно, самый распространенный вид измерения. Невелика и частота сокращений сердца—18-30 ударов в минуту, причем она сильно зависит от температуры: при низкой температуре у рыб, зимующих на ямах, она уменьшается до 1-2; у рыб, переносящих вмерзание в лед, пульсация сердца на этот период прекращается.
Ритм сердца весьма чувствителен к изменениям температуры тела и потребности в кислороде. У большинства пойкилотермных животных он ускоряется в 2-3 раза при повышении температуры на 100С.
Минутный объем сердца – это объем крови, выбрасываемый за 1 минуту желудочком одноканального сердца.
Один из способов измерения минутного объема – определения поглощения кислорода по разности в его содержании между артериальной кровью и кровью, направляющиеся к дыхательным поверхностям (метод Фика).
| Минутный объем (л/мин)= | Количество поглощенной О2 в 1 мин. |
| Артериалоьно-венозная разница по О2 | |
Сердечный индекс – отношение минутного объема к площади поверхности тела или весу, (в степени 2/3). Минутный объем, выражают в литрах крови в 1 минуту и относят к весу тела в килограммах. Минутный объем может измениться вследствие изменения ударного объема сердца или частоты его сокращений. Минутный объем сердца у пластиножаберных больше, чем у костистых. Минутный объем составляет 18,3 мл/кг, а у акулы он достигал 22 мл/кг.
Работа сердца определяется из уравнения:
Работа = Давление в желудках * объем /с
Сердечная мышца, как и другие поперечнополосатые мышцы обладают свойствами возбудимости, проводимости, сокращения. В отличии от скелетных мышц, сердечная мышца обладает своиствами автоматического сокращения, как результат внутренних «спонтанных» процессов.
Автоматия сердца – это способность сердца ритмически сокращаться под влиянием импульсов, зарождающихся в самой сердечной мышце. У костистых рыб центры автоматии распологаются в ушковом канале, а у хрящевых рыб в венозном синусе сердца. В ушковом канале имеется мышечная ткань, выполняющая функцию проводящей системы.
Возникновение и проведение возбуждения. У позвоночных сокращение сердца имеет многогенную природу. У взрослой лягушки или рыбы импульс к нормальному сокращению сердца возникает в венозном синусе или в узле Кис-Фляка. Волна возбуждения, которая возникает в синусе, охватывает мышцу желудочков со скоростью 5-6 м/с распространяется по специализированным проводящим волокнам Пуркинье.
 Ритм сокращения и электрической активности сердца регистрируется с помощью электродов, наложенных непосредственно на поверхность тела в области сердца, называется электрографией, а полученная амплитуда и крутизна кривой электрической волны, проходящей по сердцу, электрокардиограммой (ЭКГ). Рис.28. У рыб зубцу Р предшествуют формирующийся в области венного синуса зубец V, а зубец QRST- сокращение желудочка.
Ритм сокращения и электрической активности сердца регистрируется с помощью электродов, наложенных непосредственно на поверхность тела в области сердца, называется электрографией, а полученная амплитуда и крутизна кривой электрической волны, проходящей по сердцу, электрокардиограммой (ЭКГ). Рис.28. У рыб зубцу Р предшествуют формирующийся в области венного синуса зубец V, а зубец QRST- сокращение желудочка.
Рис. 27. Электрокардиаграмма:
В гистологическом строении, сердце позвоночных состоит из ветвистых поперечно исчерченных волокон. Волокна Пуркинье в желудочках млекопитающих представляют собой мышечные волокна, приспособленные для быстрого проведения возбуждения (2 м/с). Для волокон Пуркинье в сердце характерен потенциал покоя, равный 90 мВ, и потенциал действия –120 мВ; длительность его достигает 300-500 мс. Снижение содержания Na в питательной среде ведет к падению овермута, а при его снижении на 70% проведение возбуждения прекращается.
Работа сердца сопровождается механическими (сердечный толчок) и звуковыми (сердечные тоны) явлениями. Сердечный толчок и сердечные тоны можно прослушать при помощи стетоскопа, фонендоскопа, или уловить эхограммой.
3.1.4 Регуляция работы сердца. У рыб симпатические нервы сердца часто проходят в том же стволе, что и парасимпатическое. У опистогнатовых рыб ритм сокращения сердца под действием ацетилхолина замедляется; у этих рыб обнаружена тормозная холинэргическая вагусная иннервация венозного синуса и предсердия. Вагусное торможение сердца у пластиножаберных (Scyllium) может быть достигнуто рефлекторного при афферентном раздражении блуждающего и поджаберного нерва и нерва боковой линии, а у гоноидных (Acipenser) раздражение кожи, особенно на голове, плавниках и усиках, приводит к остановке сердца. У рыбы в составе блуждающего нерва, наряду с холинэргическим, т.е. тормозными волокнами идут адренэрические - стимулирующие. У некоторых рыб и амфибий, обладающие возбуждающим действием адренэргические волокна идут в составе блуждающих нервов, а у млекопитающих, птиц и рептилий в составе обособленного симпатического ствола.
Лимфатические сердца амфибий и млекопитающих рыб (угорь) обычно регулируются спинным мозгом. Перерезка нервных связей со спинным мозгом приводит к остановке лимфатического узла.
У рыб и круглоротых сердце является по сути жаберным, так как оно прогоняет кровь через сосуды жабр, после чего она по спинной аорте направляется к периферии.
У радужной форели в покое давление в брюшной аорте составляет 40/32 мм.рт.ст., а в спинной аорте –29/25 мм.рт.ст. Во время движения венозное давление достигает 9 мм.рт.ст., давление в брюшной аорте возрастает на 40%, а в спинной аорте на 16%. У форели во время движения ритм сердца ускоряется; возврату крови способствуют насосная функция хвостовой мускулатуры.
У рыб сердце иннервируется блуждающим нервом, а периферические сосуды имеют симпатическую иннервацию. Если ток воды через жабры ската или акулы прекращается, то сердце останавливается и давление падает.
Рецепторы жабр способны реагировать на такие раздражения, как кратковременное прикосновение или длительное надавливание. Псевдобронхи некоторых костистых рыб снабжены барорецепторами, афференты которых идут в составе языкоглоточных нервов.
У костистых рыб в регуляции периферических сосудов участвуют и холинэргические и адренэргические механизмы. Ацетилхолин вызывает у костистых рыб сужение жаберных сосудов, а адреналин их расширение.
Регуляция кровяного давления и кровотока у рыб, у которых кровь проходит через сердце только один раз за весь кругооборот, слагается из регуляции кровообращения в двух включенных последовательно капиллярных сетях – жаберной и периферической.
3.1.5 Строение кровеносных сосудов. Кровеносные сосуды представляют собой систему эластических трубок. Они служат для проведения крови от сердца на периферию и обратно – с периферии в сердце. Сосуды - это очень подвижные в своей организации органы: на протяжении всей жизни рыб они способны перестраиваться и развиваться вновь в связи с изменениями гемодинамических условий, тока крови, а также в связи с потребностиями организма, например при изменении или повышении функции тех или иных органов.
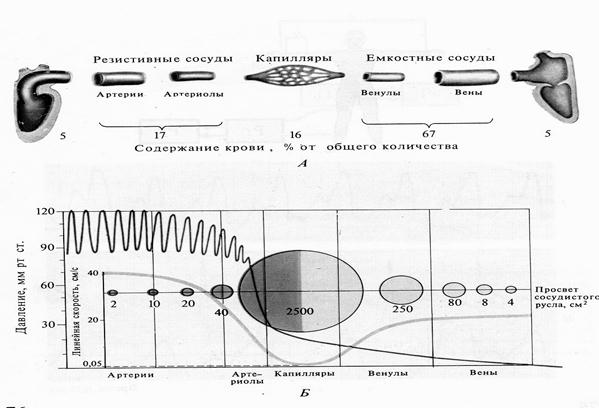
Рис. 28. Показатели гемодинамики в различных отделах сосудистого русла. А- распределение крови; Б- уровень кровяного давления, суммарный просвет сосудов и линейная скорость кровотока (у человека).
Среди кровеносных сосудов различают артерии, выносящие кровь из сердца, вены, приносящие кровь обратно в сердце, и капилляры – тончайшие волосные сосуды, соединяющие периферические концы артерий и вен друг с другом. Все эти три типа сосудов находятся в различных гемодинамических условиях и поэтому устроены неодинаково.Рис(29).
Общим для всех сосудов, равно как и для сердца, является эндотелий, который выстилает сосуды изнутри и состоит из одного слоя плоских клеток. Он изолирует кровь от непосредственного соприкосновения с любыми тканями тела. Снаружи от эндотелия (за исключением капилляров) располагаются добавочные оболочки: интима, медиа и адвентиция.
Интима, или внутренняя оболочка, представляет собой бесструктурную эластическую оболочку, которая хорошо видна на гистологических препаратах в силу своей складчатости, обусловленной посмертным уменьшением диаметра артерии. В более крупных сосудах между эндотелием и эластической оболочкой располагаются продольно идущие эластические и коллагеновые волокна.
Медиа, или средняя оболочка, является самой толстой в артериях. На ней как раз и отражаются изменения гемодинамических условий, в которых находится протекающая в сосудах кровь. По строению средней оболочки все артерии подразделяются на три типа: эластического, смешанного и мышечного строения.
В артериях эластического типа медиа построена в основном из циркулярно идущих эластических волокон и эластических окончатых мембран, с очень небольшим количеством гладких мышечных волокон. Артерии такого строения могут выдерживать очень большое давление (до 3 – 4 атмосфер) под давлением крови они легко расширяются и, благодаря своей эластичности, пассивно возвращаются в исходное положение. К числу таких артерий относятся аорта, легочная артерия и наиболее крупные ближайшие к сердцу артерии.
В артериях другого крайнего, мышечного типа медиа состоит исключительно из циркулярно идущих гладких мышечных волокон, расположенных в один или во много слоев. Такие артерии уже способны активно уменьшать свой диаметр - сокращаться, тем самым активно помогать работе сердца. Поэтому мускулатуру артерий в целом рассматривают как «периферическое сердце». К артериям мышечного типа относится большинство артерий среднего и мелкого диаметров.
Артерии смешанного, или мышечно – эластического, типа характеризуются наличием в медиа как эластических волокон, так и гладких мышечных клеток. При этом в артериях, ближайших к сердцу, преобладают эластические волокна, а в более отдаленных – мышечные волокна. К числу артерий такого типа относятся, например, подключичные и сонные артерии.
Адвентиция, или наружная оболочка, состоит из рыхлой соединительной ткани с продольными эластическими и коллагеновыми волокнами; иногда она содержит гладкие мышечные волокна. Она соединяет сосуды с окружающими органами и тканями. В мелких сосудах она почти неразличима. В организме артерии находятся в растегнутом состоянии. Но так как в стенках их имеются продольно идущие эластические волокна и гладкие мышечные волокна, то при разрезе концы артерий уходят в глубь тканей раны.
В венах, так же как и в артериях, имеются три оболочки: интима, медиа и адвентиция. Однако, в силу совершенно иных гемодинамических условий, давление крови в венах очень низкое, а в некоторых даже отрицательное. Оболочки в венах, за исключением адвентиции, развиты очень слабо, в особенности медиа, которая почти отсутствует. Поэтому стенки вен, по сравнению с артериями, очень тонкие, а диаметр вен значительно крупнее.
Характерной особенностью вен являются кармашковые клапаны, образованные складками интимы вместе с эндотелием. Таких клапанов особенно много в венах, по которым кровь течет против своего веса, например, в венах конечностей.
Добавочные оболочки артерий и вен имеют свои собственные кровеносные сосуды – «сосуды сосудов» - и нервы.
Капилляры – это важнейшие кровеносные сосуды. Стенка их состоит из одного только эндотелия. Снаружи от эндотелия располагаются отдельные клетки Руже (они же перициты, или адвентицинальные клетки). Этим именно клеткам приписывают функцию сокращения просвета капилляров, однако не исключена возможность, что эта функция принадлежит и эндотелию.
Капилляры обычно образуют более или менее густые сети, вставленные между концевыми ветвлениями артерий и началом вен, т.е. между артериолами и венулами. Диаметр капилляров очень различен (от 4 до 50 м), но он не бывает меньше диаметра эритроцита, свойственного для данного вида животного. Наиболее крупные капилляры встречаются в печени, в костном мозге, в зубной пульпе; наиболее мелкие – в мозге, в мышцах и в некоторых других органах.
Количество капилляров зависит от активности органа; чем активнее орган, тем капилляров больше, и наоборот. Общее количество капилляров в теле огромное. Их общая длина у человека определяется в 100 000 км, а поверхность в 6 000м2. При замкнутой системе кровообращения кровь не вступает в контакт с клетками. Внутренней средой для клеток является тканевая жидкость, которая обменивается с кровью газами, питательными веществами и метаболитами через стенки капилляров.
Ход и ветвление сосудов. Как правило, артерии идут вместе с нервами, образуя сосудисто – нервные пучки. Последние располагаются между мускулами, а к внутренним органам они направляются между листками серозных оболочек, в брыжейках и в связках. От спинной аорты идут артерии к внутренним органам и мускулатуре. Все крупные сосуды идут магистралями, кратчайшим путем и носят различные названия, так например брюшная аорта в брюшной полости. По пути следования магистрали отдают ветви – артерии ко всем органам, мимо которых они проходят, например-приносящие жаберные артерии.
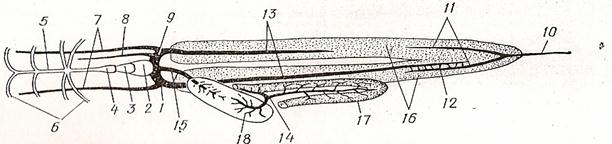
Рис.29 Схема кровеносной системы костистой рыбы (вид снизу):
1-венозная пазуха, 2-предсердие, 3-желудочек, 4- луковица аорты, 5-брюшная аорта, 6- приносящие жаберные артерии, 7- передние кардиальные вены, 8- яремная вена, 9-кювьеров проток, 10-хвостовая вена, 11-воротные вены почек, 12- анастамозы между воротной веной правой почки и правой задней кардиальной веной,13- задние кардиальные вены, 14 - воротная вена печени, 15 – печеночная вена, 16 - почки, 17 - кишечник, 18 – печень черным показаны сосуды с венозной кровью, белым с артериальной.
Во всех органах и тканях артерии распадаются на капилляры. Собирающие венозную кровь, венозные капилляры впадают в вену, несущую кровь к сердцу. Хвостовая вена, начинающаяся в хвостовом отделе, войдя в полость тела, разделяется на воротные вены почек. В почках разветвления воротных вен образуют воротную систему, а выйдя из них, сливаются в парные задние кардинальные вены. В результате слияния вен задних кардинальных с передними кардинальными (яремными), собирающими кровь из головы, и подключичными, приносящими кровь из грудных плавников, образуются два Кювьерова протока, по которым кровь попадает в венозный синус.
Кровь из пищеварительного тракта (желудка, кишечника) и селезенки, идущая по нескольким венам, собирается в воротную вену печени, разветвления которой в печени образуют воротную систему. Собирающая кровь из печени печеночная вена впадает прямо в венозный синус. Рис.29
Контрольные вопросы.
1.Круги кровообращения,механизм движения крови по сосудам.
2.Анатомо –морфологические особенности строения сердца рыб.
3.Морфологические и функциональные особенности сердечной мышцы.
4.Что такое сердечный цикл,его периоды и фазы?
5.Проводящая система сердца, механизм распространения возбуждения.
6.Биоэлектрические явления в сердце и механизм формирования ЭКГ.
7.Какик имеются функциональные типы кровеносных сосудов, и их роль в кровообращении?
8.Механизм регуляции работы сердца, сосудов.
9Дайте определение капиллярам.
10.Морфологические особенности строении артерии и вен
ГЛАВА 4
4.1 ДЫХАТЕЛЬНАЯ СИСТЕМА И ГАЗООБМЕН. Все растения и животные имеют ряд ферментов анаэробного обмена, однако большую часть энергии они получают в результате окислительных процессов. Окислительные процессы, обеспечивающие энергией все функции организма животных, связаны с постоянным притоком О2 и выносом СО2, образующегося в результате окисления различных веществ. Различают внутренний и внешний газообмен (внутреннее и внешнее дыхание): в первом случае имеется в виду обмен газов между клетками и кровью, во втором - между кровью и внешней средой.
4.1.1 Содержание кислорода в окружающей среде. Кислород, поступающий через дыхательную поверхность в организм или же прямо в клетки, должен диффундировать через водный раствор. Возможность использования кислорода зависит от его концентрации в воде, непосредственно окружающий организм. Максимальная концентрация кислорода определяется его содержанием в атмосфере (20,95% в сухом воздухе); парциональное давление кислорода в воздухе составляет на уровне море 159 мм.рт.ст. Содержание двуокиси углерода в сухом воздухе обычно составляет 0,03% (при обычных условиях это соответствует парциональному давлению 0,228 мм.рт.ст.); в промышленных районах концентрация СО2 значительно выше.
Количество О2 и СО2 в воде можно выражать, как концентрацию (в миллиметрах, миллиграммах или милях на литр) или как парциальное давление (в мм.рт.ст.) данной газовой фазы, уравновешенной с водой. На содержание кислорода в воде влияет и ряд экологических факторов. Так, перемешивание воды (быстрое течение, пороги и водопады, шторм, волны) повышает насыщение воды кислородом, тогда как в замкнутых стоячих водоемах наблюдается противоположная ситуация. Зеленые растения так же способствуют увеличению количества кислорода в воде. Сильно обедняются кислородом воды, содержащие большое количество органики (ил, детрит), поскольку расход кислорода на ее разложение может превышать его поступление в водоем. Это явление особенно выражено при высокой температуре и в зимний период, когда водоемы покрыты льдом и поступление кислорода практически приостанавливается.
Концентрация кислорода порядка 7-11 см3/л характерна для хорошо аэрированных водоемов; нередко эта величина может снижаться до десятых долей кубических сантиметров на 1 л воды. Для водного организма показатель уровня СО2 в среде не может превышать 5 мм.рт.ст. В естественных пресных водоемах концентрация кислорода может значительно измениться в зависимости от температуры воды, степени ее насыщенности воздухом, интенсивности фотосинтеза и потребления О2 организмами. Ниже термоклина, где вода почти не перемешивается, кислорода может быть намного меньше, чем у поверхности. В теплых поверхностных слоях воды, где идет интенсивный фотосинтез, концентрация кислорода может превысить ту, которая установилась бы при равновесии с воздухом.
Концентрация кислорода у поверхности, где много активных водорослей, весьма высока; на средних глубинах она обычно уменьшается в результате потребления О2 нефотосинтезирующими организмами, и здесь имеются слои с минимальным содержанием кислорода. Однако, в глубоководной зоне, уровень кислорода соответствует насыщению воздухом. Гидростатическое давление возрастает примерно на 1 атм. с каждыми 10м. глубины, но отношение между концентрациями растворенных газов в воде и в воздухе остается таким же, как и у поверхности, поэтому содержание их в воде (мг/л) не изменяется. Очень низкие концентрации О2 могут быть в иле, где кислород используется не только в биологических реакциях.
Кислород диффундирует в воздухе примерно в 3 млн. раз быстрее, чем в воде. При 200С в воде растворяется в 28 раз больше двуокиси углерода, чем кислорода, а коэффициент диффузии СО2 в воде в 25 раз выше, чем для кислорода.
Особенности водной среды обуславливают соответствующее строение и функции дыхательной системы. У позвоночных животных, высоким уровнем обмена веществ, простой контакт дыхательной поверхности с водной средой (кожа, пассивно свешивающиеся в воду жабры), не в состоянии обеспечить необходимую скорость поступления кислорода в организм.
Водное дыхание рыб осуществляется при помощи жабр и кожи. Основным органом дыхания являются жабры, а вспомогательным - кожа. Форма жабр разнообразна и зависит от видовой принадлежности и подвижности: мешочки со складочками (у рыбообразных), пластинки, лепестки, пучки слизистой, имеющие богатую сеть капилляров. Есть виды рыб, приспособившиеся частично и к воздушному дыханию (двоякодышащие, прыгун, змееголов и др.).
У рыб кровь в капиллярах жабр течет в направлении, противоположном току воды; этот противоточный механизм обеспечивает практически полное насыщение крови кислородом. Показатели эффективности жаберного дыхания служит доля кислорода (в %), извлекаемая из воды, проходящей через жаберную камеру. Функция жабер может регулироваться частотой и амплитудой их движений и изменениям скорости кровотока. Главной функцией жабр является газообмен - поглощение кислорода и выделение углекислого газа, жабры также участвуют в водно солевом обмене, выделяя аммиак, мочевину, поглощая и выделяя ионы солей, особенно ионы натрия.
На жаберных дугах сидят рядами тонкие жаберные лепестки, слизистая оболочка которых пронизана сетью капилляров кровеносной системы. Вода, омывающая жабры, отдает растворенный в ней кислород крови и уносит выделенный из крови углекислый газ. Некоторые виды рыб, живущие постоянно или временно в воде, бедной кислородом, приспособились к дыханию кислородом воздуха за счет изменения строения различных органов (плавательный пузырь, задний отдел кишечника, жаберные дуги).
Дыхание эмбрионов и предличинок рыб осуществляется за счет густой сети кровеносных сосудов на желточном мешке и в плавниковой складке. По мере рассасывания желточного мешка увеличивается количество кровеносных сосудов на плавниковых складках.
У хрящевых рыб, в частности, у акулы глотка пронизана пятью парами жаберных щелей, открывающихся наружу. В стенках глотки между жаберными щелями располагаются хрящевые жаберные дуги. К жаберным дугам прикрепляются кожистые межжаберные перегородки, доходящие до наружного жаберного отверстия и прикрывающие лежащую позади жаберную щель. На каждой стороне межжаберной перегородки располагаются многочисленные пластинчатые жаберные лепестки.
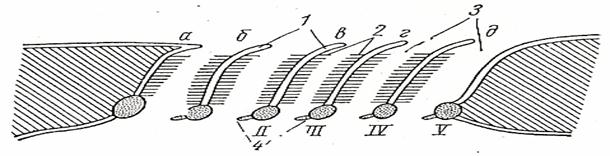 |
Рис.30. Схема жаберного аппарата акулы:
Рис. 30.Строение дыхательного аппарата акулы
1-Межжаберная перегородка, 2 – жаберные лепестки, 3- жаберная щель, 4 –жаберные тычинки,
I – V- жаберные дуги, а – первая полужабра, б,в,г,д – целые жабры.
Каждая сторона межжаберной перегородки с жаберными лепестками называется полужаберной, а две полужабры, прикрепленные к одной жаберной дуге, составляют жабру.
У колючей акулы имеется одна полужабра, расположенная на подъязычной дуге, и четыре целых жабр – на I-IV жаберных дугах; V жаберная дуга жабры не несет. Наибольшая, так называемая ложная жабра расположена на нижней стороне кожистой складки, нависающей над брызгательцем. Рис.30.
Органы дыхания у костистых рыб - жабры, имеющие эктодермальное происхождение. С каждой стороны тела располагаются четыре полные жабры; у некоторых рыб рудиментарная полужабра находится на внутренней стороне жаберной крыши, отсутствуют межжаберные перегородки. Два ряда жаберных лепестков своими основаниями прикрепляются непосредственно к костной жаберной дуге или к рудименту межжаберной перегородки, а их свободные окончания свешиваются в околожаберную полость.
У костистых рыб жаберный аппарат состоит из пяти жаберных дуг, располагающихся в жаберной полости и прикрытых жаберной крышкой. Четыре дуги на внешней выпуклой стороне имеют по два ряда жаберных лепестков, поддерживаемых опорными хрящами. Эта полость снаружи прикрыта костной жаберной крышкой, имеющей существенное значение в акте дыхания. Рис. 31
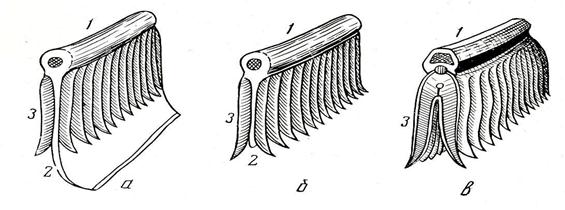
Рис. 31. Развитие жаберного аппарата (схематические отрезки):
а – хрящевой рыбы; б – химеры; в – костистой рыбы;
1 – жаберная дужка; 2 – жаберная перегородка; 3 – жаберные лепестки
С внутренней стороны каждой жаберной дуги имеются многочисленные отростки – жаберные тычинки, идущие по направлению к соседней жаберной дуге. Жаберные тычинки образуют своеобразный цедильный аппарат, препятствующий выходу пищевых частиц из глотки через жаберную полость наружу. У видов, питающихся планктоном (например, сельдей), этот аппарат представлен особенно длинными и густо сидящими тычинками. Через стенки кровеносных капилляров в жаберных лепестках, происходит газообмен крови с омывающей жабры водой, (приносящие жаберные артерии) проходит по жаберным дугам в основании жаберных лепестков. Жаберные лепестки покрыты тонкими складками — лепесточками. В них и происходит газообмен. Число лепестков варьирует; на 1 мм жаберного лепестка их приходится:у щуки — 15, камбалы — 28, окуня — 36. В результате полезная дыхательная поверхность жабр очень велика. К основанию жаберных лепестков подходит приносящая и выносящие более крупные сосуды - жаберная артерия, ее капилляры пронизывают лепесточки; из них окисленная (артериальная) кровь по выносящей жаберной артерии попадает в корень аорты. В капиллярах кровь течет в направлении, противоположном току воды. Рис.32
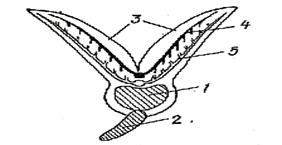
| Рис.32. Схема поперечного разреза жаберной дуги: 1-жаберная дуга, 2-жаберная тычинка, 3- жаберные лепестки, 4-приносящая жаберная артерия, 5-выносяшая жаберная артерия |
4.1.2 Газообмен в водной среде. Механизм газообмена в водной среде, заключается в диффузии газов по градиенту их концентрации.
Потребление кислорода рыбами зависит от температуры, времени года часа дня, от активности питания, его размеров, стадии жизненного цикла, а также от его снабжения кислородом и генетических особенностей.
Активность. Мышечная и другая форма активности влияет на потребление кислорода. Потребление О2 организмом в состоянии физиологического покоя намного ниже, чем при движении или физиологической нагрузки на организм. Так у рыб (форель) обмен кислорода в активном состоянии превышает в 4 раза стандартный обмен. Есть различия в интенсивности обмена и по полу: так самцы на всем протяжение жизни потребляют О2 больше чем самки.
Многие рыбы способны к эффективной регуляции дыхания. При пониженном содержание кислорода в воде степень поглощения кислорода остается неизменной, а частота дыхательных движений возрастает. С повышением температуры общее потребление О2 повышается, усиливается вентиляция, процент поглощения О2 из воды увеличивается в 2 раза. Это объясняется противоточным движением крови, при котором оттекающая из жабр кровь переходит в соприкосновении с водой богатой кислородом и бедной СО2.
Животные используют для дыхания кислород, растворенный в воде. Растворимость кислорода невелика: при 150С и давление сухого газа над водной поверхностью в 1 атм.(101,3 кПа), в 1 л воды растворяется около 34 мл О2. У рыб – как хрящевых, так и костных - вариации основного типа дыхания связаны с приспособлением к различным уровням активности и фактором окружающей среды. У очень активных рыб общая поверхность жабр (мм2 на 1г веса тела) больше, чем у малоподвижных форм, так у менхэден составляет 1241 мм2; скумбрия 1040 мм2; голец 275-432 мм2; опсанус 151-189 мм2. Так, у быстро плавающей макрели общая жаберная поверхность в пять с лишним раз больше, чем у придонной рыбы – удильщика, почти не совершающего активных движений. С помощью электронного микроскопа на пластинках жаберных нитей можно видеть микрогребни, увеличивающие рабочую поверхность жабр. У всех рыб жаберный аппарат устроен так, что вода активно прокачивается сквозь систему жаберных лепестков, через поверхность которых происходит газообмен.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 3120; Нарушение авторских прав?; Мы поможем в написании вашей работы!