КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физиология дыхания рыб 2 страница
|
|
|
|
У костных рыб движения ротового и жаберного аппаратов сочетают нагнетательный (ротовая полость) и всасывающий (жаберная полость) принципы, что обеспечивает интенсивное продвижение воды сквозь жабры.
У акуловых рыб этот механизм выражен слабее: при быстром движении, сопряженном с большими затратами энергии, для покачивания воды через жабры используется само движение: рыба плывет с открытым ртом, и вода проталкивается через жабры тем интенсивнее, чем выше скорость движения. Такой же «пассивный» темп жаберной вентиляции отмечен у тунцов. При форсированном давлении это дает значительный (до 30%) энергетический выигрыш при критических скоростях.
Во всех случаях вода проходит сквозь жабры в одном направлении; движений типа «вдох-выдох» у водных позвоночных нет, вода прокачивается через жабры практически непрерывно давление ротовой полости на протяжении всего дыхательного цикла выше, чем в жаберной.
Извлечение О2 из воды в жабрах костистых рыб усиливается и вследствие того, что направление движения потока воды сквозь жаберные лепестки и тока крови в капиллярах, проходящих по вторичным жаберным пластикам, противоположны друг другу.
Такая противоточная система способствует тому, что на протяжении всей длины жаберной пластинки сохраняется градиент концентрации О2 (и СО2) в крови и в воде, благодаря чему процесс диффузии идет непрерывно, и отходящая от жабр кровь имеет почти тот же уровень насыщения О2, что и поступающая в жабры вода. Истинные жабры распологаются между двумя «насосами» – ротовым, который связан с работой мускулатуры дна ротовой полости и жаберным, который действует за счет движений жаберной крышки. В начале цикла рот открыт, и в обеих полостях существует небольшое отрицательное давление. Когда рот закрывается, давление в ротовой полости становится положительным, вода идет через жабры и в жаберной полости тоже возникает небольшое положительное давление. Когда жаберная крышка открывается, то давление в обеих полостях падает, в ротовой полости раньше, чем в жаберной; затем жаберная крышка закрывается, а рот открывается вновь.
У многих рыб обычный ритм дыхания время от времени прерывается: в результате закрытия жаберных крышек поток воды меняет направление на обратное и происходит «откашливание», очищающее жаберную полость.
При оценке эффективности дыхания нужно учитывать степень от сигенации крови, поступающей в жабры, по следующей формуле:
| Эффективность переноса = | Количество кислорода, переходящего из воды в кровь |
| Количество кислорода в крови при max насыщении |
или точнее
| Эффективность = | Vo2 | *100 |
| Qαво2[Pio2– Puo2] |
Vo2- поглощение кислорода
Q- объем крови, проходящей через жабры за данное время
αво2 - растворимость кислорода в воде (крови)
Pio2 - парциальное давление кислорода в протекающей воде
Puo2 - парциальное давление кислорода в венозной крови, входящей в жабры.
Максимальное поглощение кислорода достигалось бы в том случае, если бы оксигенированная кровь уравнивалась по величине Ро2 с приходящей к жабрам водой. Эффективность переноса О2 у форели приближается к 100%, однако рыба извлекает из воды лишь от 11 до 30% кислорода. Эта высокая эффективность указывает на то, что кровь течет навстречу воде. У поверхности жабр течение воды остается ламинарным, поэтому каждую жаберную пластику окружает тонкий не перемешивающийся слой, в котором кислород движется очень медленно. Сопротивление переносу кислорода в этом месте составляет 80-90% общего сопротивления при переходе кислорода из воды в кровь.
Гипоксия – недостаточность кислорода, как правило, вызывает у рыб рефлекторную брадикардию, т.е. увеличение ударного объема сердца, повышение сосудистого сопротивления в жаберном русле и на переферии, а также усиление гликолиза.
У рыб во время двигательной активности значительно снижается сопротивление в переферической сосудистой сети. При гипоксии эффективность поглощения кислорода снижается.
Подобные адаптации отмечены и в связи с кислородным режимом водоемов; содержание О2, отмечается удлинение жаберных лепестков, увеличение числа жаберных пластинок и возростание суммарной дыхательной поверхности, или увеличением объема воды, пропускаемой через жаберный аппарат. При гипероксии- повышении концентрации О2 дыхание, напротив, замедляется. Обычно реакция гипервентиляции сопровождается замедлением сердечного ритма (брадикардия), что отражает некоторое снижение уровня обмена веществ – метаболизма, соответственно и потребности в О2. Следовательно, «борьба за кислород» в условиях его временной недостаточности идет по двум путям: повышения интенсивности работы газообменного аппарата и некоторого снижения затрат О2 в организме. Чувствительность рыб к Ро2 значительно выше, чем к Рсо2. У форели уже при кратковременной гипоксии вентиляция возрастает в 7 раз.
При гипоксии некоторые рыбы могут использовать кислород содержащийся в газовом плавательном пузыре. У двоякодышащих рыб легкие отходят от брюшной стороны глотки; рыба дышит легкими во время длительного пребывания вне воды. Увеличение Рсо2 в воде вызывает остановку жаберного дыхания или снижает жаберное и стимулирует легочное дыхание.
4.1.3 Факторы, влияющие на потребление кислорода. Способность крови транспортировать кислород в связанном с гемоглобином состоянии зависит от общего количества гемоглобина (и соответственно от числа эритроцитов), а также от его химических свойств, в частности от сродства гемоглобина к кислороду, определяющего способность гемоглобина к насыщению кислородом при данном его парциальном давлении. У рыб, живущих в стоячих, богатых органикой водоемах, сродство гемоглобина к кислороду выше. Гематологические отличия связаны и с подвижностью рыб: обитатели хорошо аэрированных водоемов обычно высокоподвижны. Рыбы из водоемов с низким содержанием кислорода, обычно ведут малоподвижный образ жизни при появлении признаков кислородной недостаточности происходит выброс депонированных эритроцитов в кровь, что увеличивает ее кислородную емкость.
При повышении температуры вначале возрастает объем вентиляции, а затем и частоты дыхания, ритм сердца замедляется из-за недостаточного снабжения кислородом сердечной мышцы, и тогда работа обоих дыхательных механизмов может оказаться несогласованной.
Более активные рыбы имеют большую поверхность жабр, у окуня она почти в 2,5 раза больше, чем у камбалы. Противоток крови в капиллярах и омывающей жабры воды обеспечивает полное насыщение крови кислородом. При плавании рыбы ток воды может создаваться за счет движения с открытым ртом. Таким образом, жабры расположены как бы между двумя насосами создает прокачивание воды и вентиляцию жабр. За сутки через жабры прокачивается не меньше 1 м3 воды на 1 кг массы тела.
Внешнее дыхание. Под “внешним” дыханием, или газообменом понимают способы снабжения организма кислородом и удаление из него углекислоты, а под “тканевым” же дыханием понимают совокупность ферментативных реакций окислительных и неокислительных, в результате которых накапливается энергия в форме, доступной для биологической работы.
Интенсивность обменных процессов можно определить, измерить двумя путями: а) методом прямой калориметрии, б) непрямой калориметрии, а также по количеству потребляемого кислорода, образованного тепла или выделенной двуокиси углерода.
Концентрация кислорода падает с увеличением высоты. На дне прудов и озер, морских впадинах также наблюдается недостаток кислорода.
Концентрацию кислорода выражают по-разному, в воздухе газовую смесь выражают в % или парциальном давлении; жидкости – в миллиметрах, миллиграммах или молях растворенного газа на литр, в атмосферах – мм.рт.ст. парциального давления.
4.1.4 Способы дыхания. Существуют следующие способы дыхания: прямая диффузия, кожная, кишечное, легочное, дыхание через жабры, трахейное.
При нехватке кислорода в воде рыбы всплывают на поверхность воды и дышат атмосферным воздухом путем заглатывания. Электрический угорь в норме дышит воздухом через дополнительные дыхательные органы во рту, а также и кожным дыханием. Голец Misgurnus, живущий в прудах, при нехватке О², переходят от жаберного к кишечному дыханию – за счет заглатывания воздуха.
Прямая диффузия через поверхность кожи. Все поверхности клеток проницаемы для кислорода. При отсутствии транспортной системы, кислород проникает в организм рыб путем диффузии. Все многоклеточные животные в той или иной мере обладают кожным дыханием. У угрей через кожу поступает 60%, а через жабры всего лишь 40% кислорода. Водно - кожное дыхание относится к дополнительным приспособлениям, помогающим переносить неблагоприятные кислородные условия, т. е. использование растворенного в воде кислорода при помощи кожи, и воздушное дыхание—использование воздуха при помощи плавательного пузыря, кишечника или через специальные добавочные органы. Дыхание через кожу тела - одна из характерных особенностей водных животных. По интенсивности такого дыхания пресноводных рыб делят на три группы:
1. Рыбы, приспособившиеся жить в условиях сильного дефицита кислорода. Это рыбы, населяющие хорошо прогреваемые, с повышенным содержанием органических веществ водоемы, в которых часто наблюдается недостаток кислорода. У этих рыб доля кожного дыхания в общем дыхании составляет 17- 22%, у отдельных особей—42- 80%. Это карп, карась, сом, угорь, вьюн. При этом рыбы, у которых кожа имеет наибольшее значение в дыхании, лишены чешуи или она мелкая и не образует сплошного покрова.
2. Рыбы, испытывающие меньший недостаток кислорода и реже попадающие в неблагоприятные условия. К ним относятся рыбы, обитающие у дна, но в проточной воде, осетровые — стерлядь, осетр, севрюга. Интенсивность кожного дыхания у них составляет 9-12%.
3. Рыбы, не попадающие в условия дефицита кислорода, живущие в проточных или непроточных, но чистых, богатых кислородом водах. Интенсивность кожного дыхания не превышает 3,3 - 9%. Это сиги, корюшка, окунь, ерш. Через кожу происходит также выделение углекислоты. В извлечении кислорода из воздуха во влажной атмосфере участвует не только поверхность тела, но и жабры.
Кожное дыхание для представителей обитающих в водной среде имеет важное значение. Хорошо известен своим «наземным» образом жизни илистый прыгун Periophthalmus, обитающий в болотных эстуариях тропической зоны. Эта рыба подолгу находится вне воды, передвигаясь с помощью грудных плавников. Органом воздушного газообмена у нее служит кожа.
Важное значение при этом имеет температура. Наибольшей выживаемостью во влажной среде отличаются карась (11 сут), линь (7 сут), сазан (2 сут), в то же время лещ, красноперка, уклея могут жить без воды всего несколько часов и то при низкой температуре. При перевозке живой рыбы без воды кожное дыхание почти целиком удовлетворяет потребность организма в кислороде.
Жаберное дыхание. Основная масса водных позвоночных дышат через жабры. Жабры круглоротых и рыб обычно спрятаны, они вентилируются дыхательными движениями рта и жаберной крышки. У некоторых рыб и сами жабры обладают большой подвижностью.
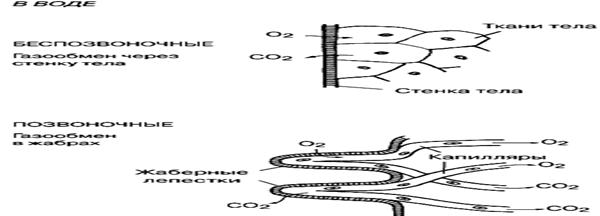 Рис. 35. Механизм газообмена у рыб А-через кожу; Б-через жабры
Рис. 35. Механизм газообмена у рыб А-через кожу; Б-через жабры
У двоякодышащей рыбы в гипоксической воде усиливается как легочное, так и жаберное дыхание. При повышении уровня СО2 в крови (гиперкапния) роль жабр в дыхании уменьшается, а дыхание воздухом усиливается. Хеморецепторы находятся на наружной стороне жабр или в выносящих жаберных сосудах. Гипоксия стимулирует вентиляцию, как легких, так и жабр; гиперкапния усиливает вентиляцию жабр в большей степени, чем легких, а при сочетании гипоксии с гиперкапнией, вентиляция легких возрастает, а жабр снижается.Использование жабр для воздушного дыхания возможно только при сохранении их поверхности во влажном состоянии.
Кишечное дыхание. У некоторых рыб, живущих в неблагоприятных условиях, выработались приспособления как, дыхание при помощи пищеварительного тракта - в различных отделах кишечника. В стенках кишечника образуются скопления капилляров. Воздух, заглатываемый ртом, проходит через кишечник, и в этих местах кровь поглощает кислород и выделяет двуокись углерода, при этом из воздуха поглощается до 50% кислорода. Такой вид дыхания свойствен вьюновым, некоторым сомовым и карповым рыбам; значение его у разных рыб неодинаково.
Воздушное дыхание рыб. Ряд видов костных рыб используют атмосферный воздух как дополнительный источник кислорода. Среди рыб с воздушным дыханием преобладают обитатели пресных вод или эстуариев тропической зоны. Данные отложения таких водоемов богаты органикой, интенсивное разложение ее при высокой температуре усиливает недостаток растворенного в воде кислорода.
Для воздушного дыхания могут использоваться жабры, слизистая ротовой и околожаберной полостей, кишечник, плавательный пузырь, кожа. Извлеченный из воды угорь вначале использует кислород, а затем начинает получать все больше кислорода через кожу, кроме того, дышит воздухом, проходящим через жабры. На воздухе жабры обеспечивают лишь одну треть, а в воде от 85 до 90% общего поглощения кислорода. При переходе из воды на воздух интенсивность метаболизма уменьшается вдвое и ритм сердца замедляется. У костистых рыб (линя, плотвы, форели) жабры расположены между ротовой и бронхиальной полостями: повышение давления в ротовой полости предшествует слабому насасыванию воды в бронхиальной полости. Этот насасывающий механизм хорошо развит у донных рыб. Такое чередование двух “насосов” имеется у акул и скатов.
Многие виды используют для воздушного дыхания плавательный пузырь, примеры такого типа газообмена – панцирная щука. У панцирной щуки путем воздушного дыхания в организм поступает 70-80% О2.
Настоящие легкие свойственны африканскому многоперу и двоякодышащим рыбам (Dipnoi), которые заселяют стоячие водоемы, подверженные регулярному пересыханию.
Другим способом использования атмосферного воздуха служит образование специальных добавочных органов дыхания как у лабиринтовых рыб, лабиринт - расширенный карманообразный участок жаберной полости, складчатые стенки которого пронизаны густой сетью капилляров, в которых происходит газообмен. Таким способом рыбы дышат кислородом атмосферы и могут находиться вне воды в течение нескольких дней (тропический окунь-ползун Anabas sp. выходит из воды и лазит по камням и деревьям).
4.1.5 Плавательный пузырь и газообмен Рыбы обладают специальными приспособлениями, чтобы держаться в толще воды. Основным органом, рыб регулирующих свой удельный вес, а также приуроченность к определенным слоям воды, является плавательный пузырь. Лишь немногие рыбы, обитающие в толще воды, его не имеют, это акулы и некоторых скумбриевых. Эти рыбы регулируют свое положение в том или ином слое воды только при помощи движения плавников.
Существуют два типа газовых пузырей: открытые у открытопузырных рыб и закрытые у закрытопузырных. Открытопузырные могут заглатывать в пузырь воздух; кроме того, и те и другие рыбы обладают способностью выделять в полость пузыря газ. Газовый (плавательный) пузырь выполняет в первую очередь гидростатическую роль, и регулируют глубину погружения. Так глубоководные рыбы всплывают или погружаются в зависимости от объема газа в пузыре.
Вторая функция плавательного пузыря – повышение чувствительности сенсорных органов: пузырь усиливает звуковые волны, передаваемые водой. У некоторых рыб плавательный пузырь служит для испускания звуков. Кроме того, плавательный пузырь может играть роль поставщика кислорода при респираторном стрессе, как, например, у данных рыб. Плавательные пузыри более широко распространены у поверхностных и пелагических рыб, обитающих на глубине до 600 м., чем у донных.
Плавательный пузырь служит не только для изменения удельного веса рыбы, он играет роль и органа, определяющего величину наружного давления у ряда рыб. У большинства вьюновых – Cobitidae, ведущих данный образ жизни, плавательный пузырь сильно редуцирован, и его функция как органа, воспринимающего изменения давления, является основной. Рыбы могут воспринимать даже незначительные изменения давления, их поведение меняется при перемене атмосферного давления. В Японии некоторых рыб специально содержат для этой цели в аквариумах и по изменению их поведения судят о предстоящей смене погоды.
Для поддержания плавучести в морской воде нужен плавательный пузырь, объем которого составлял бы 3-5% объема тела, а в пресной воде - несколько больше. Закрытопузырные рыбы наполняют пузырь газом исключительно путем секреции, открытопузырные же могут заглатывать часть газа, но при отсутствии доступа к поверхности воды они способны секретировать его. Плавучесть рыб, лишенных плавательного пузыря, часто достигается за счет увеличения собственного веса, в частности жирности. В особенности увеличивается вес печени, что составляет 23% от веса тела, а ее удельный вес- 0,94.
У большинства более древних групп рыб плавательный пузырь соединен с кишечником при помощи специального протока. У остальных рыб – окунеобразных, трескообразных и других костистых, с возрастом связь плавательного пузыря с кишечником не сохраняется. Плавательные пузыри у некоторых сельдевых и анчоусов, у океанической сельди, шпроты, хамсы имеют два отверстия. У рыб, имеющих соединение плавательного пузыря с кишечником, поступление и выделение газа из плавательного пузыря происходит в значительной степени через ductus pneumaticus, это наружное отверстие в задней части пузыря, открывающееся непосредственно за анальным отверстием. Это отверстие позволяет рыбе за короткий срок удалить из плавательного пузыря лишний газ. При этом у опускающейся на глубину рыбы излишний газ появляется в пузыре под воздействием возрастающего по мере погружения рыбы давления воды на ее организм.
За исключением некоторых сельдевых все рыбы, обладающие плавательным пузырем, не могут быстро переходить из поверхностных слоев на глубины и обратно, а закрытопузырные рыбы могут перемешаться в слое воды. В связи с этим у большинства видов, совершающих быстрые вертикальные перемещения, плавательный пузырь либо совершенно отсутствует, либо редуцирован, а удержание в толще воды осуществляется за счет мускульных движений тела или плавников. Редуцируется плавательный пузырь и у многих донных рыб, многих бычков, морских собачек, вьюновых и др. Редукция пузыря у донных рыб, связана с необходимостью обеспечить больший удельный вес тела.
Наполнение газом плавательного пузыря происходит не сразу по выходе из икринки. Пока выведшиеся свободные эмбрионы проходят стадию покоя, подвесившись к стеблям растений или лежа на дне, газа в плавательном пузыре у них нет. Наполнение плавательного пузыря газом у одних видов осуществляется за счет наружного воздуха, у других газ выделяется крупными клетками, расположенными внутри плавательного пузыря. У многих рыб проток, соединяющий кишечник с пузырем, во взрослом состоянии отсутствует, а у их личинок он имеется.
Секреция газа в плавательный пузырь. Некоторые водные животные способны накачивать газы в пузырях или газовых камерах против значительного градиента давления. Наиболее изучен механизм данного процесса секреции газов в плавательный пузырь у костистых рыб. У закрытопузырных рыб весь газ плавательного пузыря происходит из газовой железы. Наибольшее количество газа секретируется у открытопузырных.
У большинства рыб, обладающих плавательным пузырем, имеются механизмы, как для секреции газа, так и для его поглощения. В регулировании объема пузыря могут участвовать железы и мышцы. Мышцы, ответственные за наполнение пузыря воздухом, снабжены нервными холинэргическими, а их антагонисты – адренэргическими окончаниями. Существующие медиаторы действуют как на мышцы, так и на железы.
У рыб с замкнутым плавательным пузырем после первоначального наполнения газом извне дальнейшее изменение количества и состава газа регулируется путем его выделения и поглощения кровью. У таких рыб на внутренней стенке пузыря находится красное тело – образование, чрезвычайно густо пронизанное кровеносными капиллярами. Так, в двух красных телах плавательного пузыря угря имеется 88000 венозных и 116000 артериальных капилляров (в красных) общей длиной 352 и 464м. В то же время объем всех капилляров в красных телах угря составляет всего лишь 64мм 3. По величине красное тело варьирует у рыб от небольшого пятна до мощной газоотделительной железы, состоящей из цилиндрического железистого эпителия. Скорость наполнения газом плавательного пузыря за счет газов крови различна, так рыбе с открытым пузырем для наполнения плавательного пузыря требуется 5-7 дней, а угрю 12-20 ч.
Газ, выделяемый в плавательный пузырь, сильно отличается по составу от воздуха. На глубине, где действует гидростатическое давление, количество растворенного газа возрастает на 14% при каждом увеличении давления на 100 атм., так что на глубине 1000 м, Ро2 достигает 0,228 атм. Газ в плавательном пузыре глубоководной рыбы может содержать 65-95% О2. Таким образом, общее давление кислорода в плавательном пузыре рыбы на глубине 1000 м составляет около 90 атм, т.е. активный перенос кислорода в плавательный пузырь увеличивает давление О2 в 394 раза.
Состав газа в плавательном пузыре у морских рыб, взятых на глубине 100 м составил: так, количество азота соответствует 0,8% давления воздуха, или 0,8 атмосферного давления, на глубине 900 м., парциальное давление азота в плавательном пузыре колебалось от 5 до 15 атм. Когда на поверхности воды напряжение азота соответствует 80% давления воздуха - 0,8 атмосфер. На каждые 10м. глубины давление повышается приблизительно на 1 атм. Следовательно, у рыбы на глубине 100 м. при выделение газа должен преодалеваться градиент давления в 10 атм. Если рыба всплывает, расширение газа может вызвать разрыв плавательного пузыря.
Состав газа в плавательном пузыре неодинаков, как у различных видов рыб, так и у разных особей одного и того же вида. В плавательном пузыре морских рыб кислорода содержится больше, чем у пресноводных.
Каждому виду свойствен свой кислородный порог, т. е. минимальная концентрация кислорода, при которой рыба гибнет. Форель начинает задыхаться при концентрации кислорода 1,9 мг/л, судак и лещ погибают при 1,2, плотва и красноперка — при 0,25 - 0,3 мг/л; у сеголетков-карпов, выращенных на естественной пище, кислородный порог отмечен при 0,07- 0,25 мг/л, а для двухлетков—0,01-0,03 мг/л кислорода. Караси и ротаны — частичные анаэробы — несколько суток могут жить совсем без кислорода, но при низкой температуре. Предполагают, что сначала организм использует кислород из плавательного пузыря, затем — гликоген печени и мышц.
Подвижные и хищные рыбы имеют большой запас кислорода в плавательном пузыре, который расходуется организмом при бросках за добычей, когда поступление кислорода через органы дыхания оказывается недостаточным. В неблагоприятных кислородных условиях воздух плавательного пузыря у рыб используется для дыхания. Вне воды угорь использует для дыхания кислород плавательного пузыря и воздух, проходящий через кожу и жабры. Это позволяет ему даже переползать из одного водоема в другой. Осваивая различные водоемы, рыбы приспособились к жизни при разных газовых режимах. Наиболее требовательны к содержанию кислорода в воде лососевые, которым для нормальной жизнедеятельности нужна концентрация кислорода 4,4-7 мг/л; хариус, голавль, налим хорошо себя чувствуют при содержании не менее 3,1 мг/л; карповым обычно достаточно 1,9- 2,5 мг/л.
На рыб губительно действует чрезмерное перенасыщение кислородом воды. Так, летальной границей для эмбрионов щуки является 400% насыщения воды кислородом, при 350 - 430% насыщения нарушается двигательная активность эмбрионов плотвы. Прирост осетровых снижается при 430% насыщении кислородом.
Инкубация икры в перенасыщенной кислородом воде приводит к замедлению развития эмбрионов, сильному увеличению отхода и количества уродов и даже гибели. У рыб появляются пузырьки газа на жабрах, под кожей, в кровеносных сосудах, органах, а затем наступают судороги и смерть. Это называется газовая эмболия или газопузырьковая болезнь. Для поддержания оптимальной концентрации кислорода в воде, обеспечивающей наиболее эффективное течение физиологических процессов в организме рыб, нужно использовать аэрационные установки. У них повышается обмен и как результат увеличивается потребление корма и снижается кормовой коэффициент, развитие эмбрионов ускоряется, отходы снижаются.
Наибольшего развития как орган дыхания плавательный пузырь достигает у двоякодышащих рыб. У них он ячеистый и функционирует, как легкое. При этом возникает “легочный круг” кровообращения. Благодаря переплетению сосудов со встречными потоками крови кислород может переходить из вены, идущий от плавательного пузыря, непосредственно в артерию, идущую от сердца, а затем из общей вены в артерию плавательного пузыря. Таким образом, в ткани секретирующей железы может создаваться высокая концентрация кислорода. В артерии плавательного пузыря имеет место «положительный» эффект Рута, т.е. увеличение максимального уровня насыщения гемоглобина.
У некоторых глубоководных рыб полость плавательного пузыря содержит большое количество липидного материала, который состоит из фосфолипида и холестерина в соотношении 1:1. Газовая железа глубоководных рыб синтезирует холестерин. Так как кислород, возможно, хорошо растворим в этом холестеринсодержащим липиде, в пузыре может оставаться очень мало свободного газа. Напротив, у рыб, обитающих на небольших глубинах, имеется лишь тонкий слой липида на внутренней поверхности пузыря.
Сифонофоры как физалия выделяют газ в объемистый пузырь-пневмотофор. Секретируя газ, она всплывает на поверхность, а выпуская его – погружается под воду. Находящийся в пузыре газ содержит 74,4% N2, 8,9% СО, 14,4% О2, 1,1% аргона и 0,4% СО2. У основания пузыря расположена газовая железа, сходная по своей ультраструктуре с железой рыб. Формы у этой сети и железы у разных рыб разная, но принцип работы одинаковый; артерии и вены в железе распадаются на множество капилляров, а затем вновь собираются в один сосуд. Капилляры по длине достигают до 25 мм, тогда как в мышцах длина равна 0,5 мм. Газовая железа в пищеварительном пузыре выполняет также роль умножителя благодаря своему морфологическому строению, именно здесь, наблюдается высокое парциальное давление газов. У глубоководных рыб благодаря чудесной сети количество кислорода находится на нужном уровне из-за его непрерывного и диффузного физического поступления в артериальные капиляры.
Интенсивность дыхания рыб зависит от размера, возраста, подвижности, активности питания, пола, степени зрелости гонад, физико-химических факторов среды. По мере роста рыб активность окислительных процессов в тканях уменьшается, а созревание гонад, наоборот, вызывает увеличение потребления кислорода. Расход кислорода в организме самцов выше, чем у самок.
4.1.6.Регуляция процессов дыхания. Физиологические механизмы регуляции ритма дыхания у рыб изучены еще недостаточно. Ритм дыхательных движений поддерживается импульсами, идущими от клеток дыхательного центра, расположенных в продолговатом мозге. Ритмические разряды нервных импульсов в этой области можно обнаружить даже после обездвиживания рыбы тубокурарином или сукцинилхолином, а также после перерезки всех чувствительных нервных волокон, идущих к этой части мозга. Локализация кислородных рецепторов точно не выяснена. Скорее всего, они расположены не в самих жабрах (брадикардия возникает и при отключении тока крови через жабры), а на их поверхности, в спинной аорте, в ротовой полости или же в ложножабрах.
У рыб есть как периферические, так и центральные рецепторы воспринимающие уровень О2. Недостаток кислорода увеличивает как частоту, так и глубину дыхания, поэтому объем вентиляции возрастает. Избыток СО2 иногда может вызывать небольшое снижение частоты и углубление дыхания. Угорь Anguilla и форель Salmo поглощают около 80% - О2, растворенного в воде, насыщенный воздухом; при более низком содержании О2 в воде они поглощают меньше кислорода. Если уровень О2 уменьшается до 4 мл/л., то угорь и форель усиливают в 5 раз интенсивность дыхательных движений. Четыре – пяти кратное увеличение активности приводит к тому, что поглощение О2 из воды повышается на 70%, в результате рефлекторного воздействия накопившегося углекислоты.
Наиболее тонкая регуляция дыхательного процесса и работы дыхательного центра, расположенного в продолговатом мозге осуществляется за счет концентрации углекислоты в крови. Углекислота является крепким стимулятором дыхательного центра, обладает высокой растворимостью в воде и редко накапливается в тканях водных животных при нарушение их дыхания. Первичным возбудителем центра дыхания является недостаток О2 в воде. Дыхательный центр акульих и костных рыб, хорошо развиты, обладают автономной активностью. У акулы во всех жаберных ветвях девятого и десятого черепномозговых нервов имеются ингибиторные эфферентные волокна. У сома и карася центры регуляции находятся позади моторного ядра, у линя в нижнем краю фасциальных долей продолговатого мозга.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 12317; Нарушение авторских прав?; Мы поможем в написании вашей работы!