КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Понятие обмена веществ и превращения энергии
|
|
|
|
ФИЗИОЛОГИЯ ОБМЕНА ВЕЩЕСТВ
Основным процессом, характеризующим самую жизнь и непрерывно протекающим в живом организме, является обмен веществ. Этот процесс сводится к поступлению в организм из внешней среды различных веществ, их усвоению, изменению и выделению продуктов распада. При этом в организме осуществляется множество различных химических, механических, термических и электрических явлений. Непрерывно происходит превращение энергии, освобождение связанной химической потенциальной энергии и переход её в другие виды.
Обмен веществ отличает живой организм от мёртвого, органический мир от неорганического. Изменения веществ и превращения энергии осуществляются и в неорганическом мире.
Процессы обмена веществ состоят из явлений ассимиляции и диссимиляции, ассимиляцией называется сумма процессов созидания живой материи, т, е. усвоение веществ, поступающих в организм из внешней природы, образование сложных химических соединений из более простых. Ассимиляция — это происходящий в организме синтез живой протоплазмы.
Диссимиляцией называется изнашивание живой материи, разрушение, распад, расщепление входящих в состав протоплазмы веществ, в частности, белковых соединений.
Ассимиляция и диссимиляция протекают в организме одновременно и составляют единый процесс. (Рис. 43)
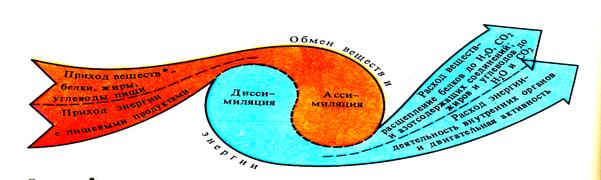
· - за исключением белков жиров,углеводов,экскрементов
Рис 43 Общее представление об обмене веществ и энергии.
Однако они не всегда взаимно уравновешиваются. В различные моменты жизни и деятельности организма один из них может преобладать.
В период роста, например, преобладают явления ассимиляции; старость характеризуется выраженным преобладанием диссимиляции.
Процессы ассимиляции и диссимиляции длительно невозможны без поступления в организм пищи и кислорода извне и выведения продуктов распада. Поэтому обмен веществ включает три этапа: 1) поступление различных органических и неорганических веществ в организм, 2) их изменение в организме и 3) выведение продуктов распада.
Источником пополнения организма веществами и энергией является пища. Пища (или корм) составляется из пищевых продуктов. Этим термином обозначается смесь пищевых (или кормовых) веществ. К пищевым продуктам (кормам) относятся, зоо- и фито - планктон. Все они содержат воду, минеральные вещества, углеводы, жиры, белки и пр. Пищевые вещества поступают в организм через пищеварительный тракт. Вместе е пищевыми органическими веществами поступает и потенциальная энергия для клеток.
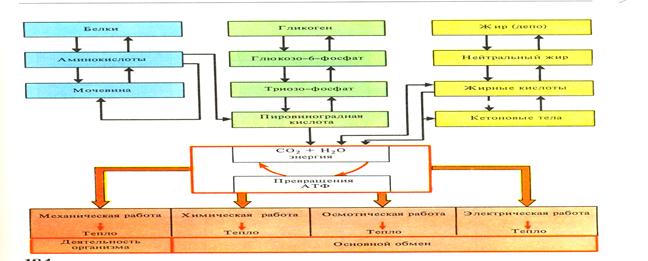
Рис.44 1) Схема превращения веществ и энергии в организме.
Характеризуя пищу с точки зрения обмена веществ и энергии, можно сказать, что пища нужна организму:
1) для построения клеток тканей организма;
2) для образования регулятора реакций - ферментов, гормонов и витаминов;
3) для пополнения организма водой;
4) как источник энергии;
5)фактор, поддерживающий на определенной высоте работу сердца, дыхательного аппарата, движение желудочно-кишечного тракта, тонус кровеносных сосудов.
Кроме того, часть пищи откладывается в организме в виде запасов (жиры, углеводы), которые необходимы организму в случае нужды, например, в период заболевания. Пища выполняет роль основной базы вещественного и энергетического обмена.
В процессе обмена веществ непрерывно происходит изменение энергии. Потенциальная (химическая) энергия органических веществ при их расщеплении освобождается и превращается в другие виды энергии в организме рыб. Обмен веществ и обмен энергии являются двумя проявлениями единого процесса. Различают обмен веществ как общий, межуточный, основной, азотистый (белковый), углеводный, жировой, минеральный и водный. (Рис. 44)
6.2 Общий обмен Под общим обменом понимают, непосредственно вещественный и энергетический обмен между внешней средой и организмом при обычных условиях жизни. Этот обмен включает рассмотрение всех этапов:
1) поступление пищи и превращение её в желудочно-кишечном тракте,
2) изменение всасывающихся веществ в клетках, тканях и органах,
3) образование и выделение продуктов распада.
6.3 Межуточный обмен — это совокупность химических превращений в клетках, тканях и органах веществ, всасывающихся из кишечника.
6.4 Основной обмен — это обмен веществ в организме находящихся в состоянии полного покоя, лишённых всяких раздражений и имеющих относительно освобождённый от пищевых масс желудочно-кишечный тракт. Основной обмен, совокупность превращений в теле, которые необходимы для поддержания основных жизненных функций. Этот обмен является характеристикой интенсивности обмена веществ и энергии. Определяющими его факторами являются рост, вес, возраст и пол. Значительная часть основного обмена у теплокровных животных падает на работу дыхательной мускулатуры (10—15%), сердца (около 4%), почек (около 5%) и печени. При нормальном течении физиологических процессов величина основного обмена довольно постоянна.
В понятия общий, основной и межуточный обмены включаются процессы превращения отдельных органических веществ, воды и минеральных веществ, т.е. белковый, или азотистый, углеводный, жировой, водный и минеральный обмен.
6.5 Белковый обмен. После расщепления в желудочно-кишечном тракте белки всасываются из кишечника в кровь в виде аминокислот и несложных полипептидов. Из них в клетках синтезируется тканевой белок. По имеющимся данным, белки плазмы могут переходить в белки тканей, в частности, печени, без предварительного распада до аминокислот.
Процесс синтеза белка в организме непрерывен, потому что там постоянно происходят разрушение клеток и распад их протоплазмы. При синтезе белков необходимо известное количество различных аминокислот. Есть аминокислоты, которые не синтезируются в организме, и недостаток их в пище ведёт к нарушению белкового обмена. К таким незаменимым аминокислотам относятся триптофан, тирозин, цистин и ряд других. Биологическое значение этих аминокислот состоит в том, что при их отсутствии или недостатке наступает отрицательный баланс белка, останавливается рост, падает вес тела.
О потребности организма в белках и о размерах его распада в организме можно судить учитывая азот, введённый в организм с пищей и выведенный из организма с мочой и калом. Этим путём устанавливается так называемый баланс азота в организме.
Среднее содержание азота в белке составляет 16%. Количество введённого белка определяется путём исследования азота, содержащегося в принятой пище. Часть воспринятого с пищей азота теряется с калом, поэтому для определения усвоенного азота вычитают количество азота в кале из количества азота в пище. Найденную величину усвоенного азота умножают на коэфициент пропорциональности — 6,25, соответственно 16% содержания азота в белке (100: 16=6,25). Таким путём узнают величину усвоенного белка в граммах. Для определения количества разрушенного белка определяют содержание азота в моче. Если азота поступает больше, чем расходуется, говорят о положительном балансе, т. е. о преобладании в организме синтеза белка над его распадом. Отрицательный баланс указывает на преобладание распада белка над его синтезом.
Задержка азота в организме, иначе ретенция его, наблюдается в период роста. С окончанием pocтa размер синтетических процессов и задержка азота резко сокращаются.
В печени может накапливаться известное количество белка, как запасный фонд, который используется организмом при недостаточном поступлении белка извне. Количество запасного белка в печени невелико. По данным исследований, значительная доля аминокислот крови поступает в тканевые элементы и также составляет там запасный фонд, особенно если эти кислоты адсорбированы в малоактивных тканях, как, например, в красных кровяных клетках, в соединительной ткани.
Одновременно с синтезом тканевого белка идёт его распад. Этот распад происходит постоянно, даже и при отсутствии поступления белка в организм. Объясняется это тем, что в результате жизнедеятельности организма всегда происходит распад некоторого количества веществ протоплазмы и разрушение некоторых клеток. Если выключить из рациона белок, а другие вещества вводить в количестве, достаточном для удовлетворения суточной потребности организма в энергии, то все же организм выделяет часть того азота, который связан с самими процессами жизнедеятельности. Эти наименьшие для организма потери белка назвали коэфициентом изнашивания.
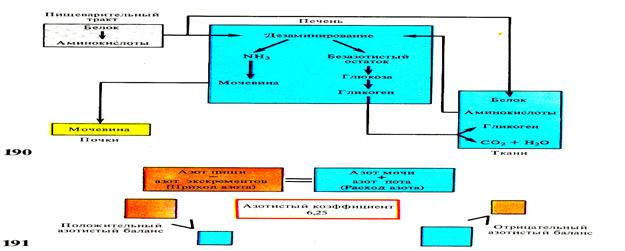
Рис.45 Схема белкового обмена, азотистый баланс
Распад белка в организме возрастает при включении его в рацион. Объясняется это, прежде всего, тем, что часть белков расходуется как энергетический материал. Освобождённый при этом азот не задерживается в организме и выделяется.
У взрослого организма количество введённого с пищей азота обычно бывает равным количеству выведенного азота. Такое состояние называется азотистым равновесием, которое служит показателем нормального белкового обмена. Организм обнаруживает сравнительно малую способность к отложению запасов белковых соединений. Поэтому при увеличенном введении белка вскоре устанавливается азотистое равновесие.(Рис. 45)
Часть белков используется на синтез в связи с распадом тканевых белков, остальное же количество служит энергетическим материалом. В результате количество усвоенных и разрушенных азотистых соединений становится равным.
Внутриклеточный распад белковой молекулы образует конечные продукты: аммиак, мочевину, мочевую кислоту и ряд других веществ, которые и выделяются с мочой. В образовании конечных продуктов белкового обмена большое значение принадлежит печени.
6.6 Обмен жиров Жиры всасываются в виде глицерина и жирных кислот. В клетках кишечника эти вещества вновь синтезируются в жировую молекулу и, чаще в виде жира данного вида животного, поступают в лимфу, а частично в кровь.
Жиры, прежде всего входят в состав клеток, как их структурная часть, в виде включений в протоплазму, и принимают участие в образовании протоплазмы, ядерного вещества и оболочки. Большое количество всосавшегося жира откладывается прозапас в жировой ткани (подкожная клетчатка, вокруг некоторых внутренних органов и внутри органов, например, в печени и мышцах).
Количество жира, входящего в структуру клеток в организме, сравнительно устойчиво, а количество запасного жира колеблется в зависимости от питания, пола, возраста, породы и конституциональных особенностей.
Жиры могут образовываться в процессе межуточного обмена из углеводов и белков. В межуточном обмене жиров большая роль принадлежит печени. Установлено, что жиры, поступающие из кишечника в кровь, задерживаются в печени, а поступающие в лимфу идут в жировую ткань подкожной клетчатки, которая составляет так называемое жировое депо организма. При усилении энергетических затрат и при голодании используется жир из депо. Существует связь между углеводами и жировым обменом в печени. При уменьшении гликогена в печени содержание в печёночных клетках жира увеличивается, и наоборот.
В процессе обмена жиры расщепляются на глицерин и жирные кислоты, которые затем подвергаются окислению. Конечными продуктами распада жира являются СО2 и Н2О.
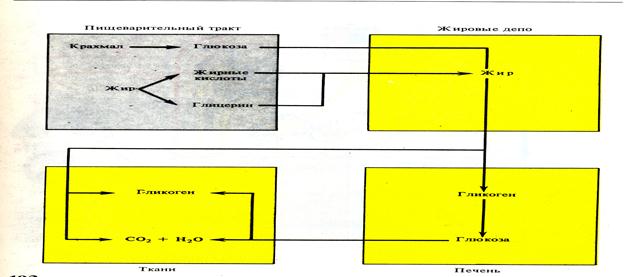
Рис.46.Схема жирового обмена (по А.Гинецинскому,1956г)
6.7 Обмен углеводов Углеводы всасываются из кишечника в кровь в виде моносахаридов, чаще в виде глюкозы (виноградного сахара). Виноградный сахар в печени синтезируется в гликоген, который составляет запасный фонд углеводов в организме. Кроме того, запасы углеводов в виде гликогена содержатся в мышцах.
Содержание сахара в крови поддерживается на определённом уровне: 0,11—0,15%. При повышении концентрации сахара выше 0,15—0,18% избыток его выделяется через почки с мочой - г л ю к о з у р и я. В нормальных условиях обмен углеводов и, в частности, содержание сахара в печени и в крови регулируются железами внутренней секреции и нервной системой. Образование углеводов может происходить в организме из белков и жиров. По мере убывания сахара в крови происходит мобилизация гликогена из мышц, благодаря чему сохраняется относительное постоянство содержания глюкозы в крови. Особенно резко увеличивается распад гликогена при мышечной работе. (Рис. 47).
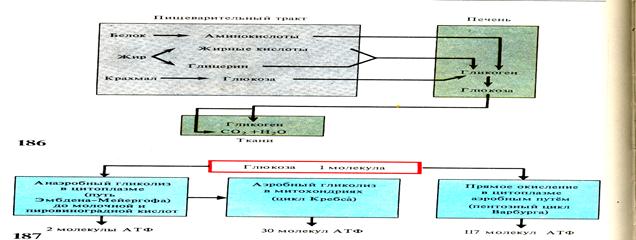
Рис.47 Схема углеводного обмена, расщепление глюкозы и образование АТФ.
Внутриклеточный распад углеводов сложен: он осуществляется сначала путём безкислородного расщепления - гликолиза, а затем путём окисления продуктов распада (молочной кислоты) до С02 и Н2О.
6.8 Обмен минеральных веществ и воды Наряду с белками, жирами и углеводами, для организма важно иметь минеральные вещества и воду. Значение их разнообразно, при их участии в организме протекают важнейшие физико-химические процессы. Благодаря минеральным солям, растворимым в воде, создаётся определённое осмотическое давление крови и тканей, сохранение которого на постоянном уровне является необходимым условием нормальной жизнедеятельности.
Минеральные вещества принимают участие в поддержании кислотно-щелочного равновесия и сохранении относительного постоянства активной реакции крови.
Вода и минеральные вещества способствуют сохранению коллоидального состояния живой протоплазмы и участвуют в построении клеточных структур. Деятельность выделительных органов, секреция пищеварительных желез и транспорт газов крови связаны с участием минеральных веществ. Значение минеральных веществ не ограничивается только их участием в физико-химических процессах. Распадаясь на ионы, они оказывают специфическое влияние на физиологические процессы. Если лишить организм воды и минеральных веществ, это приводит к тяжёлым нарушениям и смерти. Неорганические соединения в организме высших животных и человека энергетического значения не имеют. Этим они отличаются от белков, жиров и углеводов.
В обычных условиях смешанного типа питания, организм получает достаточное количество минеральных солей. Иначе обстоит дело с питанием растущего организма, который особенно нуждается в большинстве минеральных соединений.
Для развития костной и нервной ткани молоди рыб необходимо достаточное снабжение организма кальцием и фосфором. Кроме того, у растущего организма происходит задержка, натрия, калия, магния, хлора. Имеются данные, указывющие на то, что увеличение веса тела на 1 кг связано с задержкой в организме 1 г. Na.
Из кишечника минеральные вещества всасываются в виде солей и ионов. Минеральные вещества входят в состав клеток и жидкостей тела. В случае избыточного поступления солей их запас может отложиться в различных органах, а при недостатке минеральных веществ происходит мобилизация запасов. Например, кальций и фосфор откладываются в костях, железо — в печени, натрии и хлор — в коже, калий — в мышцах.
Из организма минеральные вещества выделяются с мочой, калом. С калом выделяются ионы железа, кальция и магния. Местом их выделения служит толстый отдел кишечника.
Воды в организме содержится от 35 до 63%. Содержание воды и минеральных веществ в организме подчинено регуляции через нервную систему и гуморально (через кровь).
Водный обмен организма со средой включает два противоположных процесса: поступление воды в организм и отдачу ее им во внешнюю среду. Обитатели водной среды способны получать воду через покровы или специализированные участки тканей, проницаемые для воды (например, жабры).Значение пищи в водном балансе организма заключается не только в том, что она содержит воду в тканях пищевых объектов. Усиленное питание ведет к накоплению в организме жировых запасов: в процессе окисления жиров и других органических веществ образуются так называемая метаболическая вода. Таким образом, жир в организме животных не только энергетический запас, но и внутренний источник поступления воды в клетки ткани.
Водный обмен теснейшим образом связан с обменом солей. Определенный набор солей (локов) представляет собой необходимое условие нормальной функции организма, так как соли входят в состав клеток и тканей. Играя важнейшую роль в обменных механизмах клеток, изменение количества, воды и солей нарушает осмотические процессы и ионое равновесие организма. В организме животных постоянно идет активное регулирование концентрации солевого состава жидкостей и тканей тела. Для организма водных животных важное значение имеет поддержание постоянства водно-солевого обмена, у которых осмотические процессы осуществляются не только на суборганизменном уровне, но и в зависимости от состояния окружающей водной среды.
Водно-солевой обмен у водных животных. У первично водных животных жаберные, кишечные, покровные эпителии всегда контактирует с водой, поэтому через их покровы постоянно осмотическим путем проникает вода, в том случае если концентрация солей в организме отличается от окружающей водной среды. Состав жидкостей тела рыб качественно сходен с составом морской воды. Однако количественное содержание различных ионов в теле даже морских животных может отличаться от соотношения их в окружающей их среде. Некоторые соли идут на построение скелета, а часть циркулирует в состав крови и лимфы, поддерживая постоянство внутренней среды, или, напротив, активно выводится из организма.
Ионную регуляцию в организме, связанную с поддерживанием состава и количественного соотношения различных ионов в жидкостях тела, и регуляцию осмотического давления, называют водно - солевым обменом. Изменение осмотического давления внутри организма животных приводит к изменениям давления внешней среды, таких животных называют пойкилоосмотичными (осмоконформерами). Животные, способные к активной регуляции осмотического давления поддерживают относительное постоянство внутренней среды независимо от состояния внутренней и окружающей среды, таких животных называют гомойоосмотическими (осморегуляторами).
Осморегуляция (от осмос и лат. regulo — направляю), совокупность физико-химических и физиологических процессов, обеспечивающих относительное постоянство осмотического давления (ОД) жидкостей внутренней среды (межклеточных жидкостей, лимфы, крови) у гомойосмотических животных. Механизмы осморегуляции обнаруживаются у организмов, обитающих в средах с различной концентрацией осмотически активных веществ, главным образом солей. Осморегуляция присуща всем пресноводным и наземным животным; среди морских организмов осморегуляцией обладают все позвоночные (кроме миксин) и некоторые ракообразные.
В основе физиологических механизмов осморегуляции лежит соответствующий рефлекс: изменение осмотического давления крови или межклеточной жидкости, которые воспринимаются специальными нервными клетками - осморецепторами. Возникающие в них импульсы передаются в нервные центры головного мозга, которые регулируют потребление воды и солей и их выделение осморегулирующими органами — нефридиями, почками, солевыми железами и др.
Механизм осморегуляции может быть гиперосмотическим, когда осмотическое давление внутренней среды организма больше, чем в жидкости внешней среды, и гипоосмотическим, когда внутреннее осмотическое давление меньше внешнего. В первом случае избыток воды выделяется животным главным образом через почки (растением — преимущественно через устьица); во втором — пассивную потерю воды через покровы тела организм восполняет, потребляя богатую солями воду и выделяя избыток солей в основном через солевые железы.
Пресноводные животные и морские хрящевые рыбы характеризуются гиперосмотической регуляцией. У акул и скатов это обусловлено высокой концентрацией в крови мочевины. В тело этих животных через проницаемые для воды участки покровов тела по осмотическому градиенту поступает вода. Избыток её у всех животных выделяется почками или их аналогами (сократительные вакуоли простейших, нефридии), а соли всасываются из пресной воды клетками жаберного аппарата или кожи (у земноводных).
Гипоосмотическая регуляция характерна для морских костистых рыб, морских пресмыкающихся и некоторых других. Эти животные теряют воду через покровы и с мочой. Для компенсации потерь они пьют морскую воду, опресняемую клетками солевых желёз и др. органами, выделяющими концентрированные растворы солей.
У млекопитающих основной орган осморегуляции - почки, способные выделять гипотоническую мочу при избытке воды и осмотически концентрированную — при её дефиците. Проходные рыбы (например, лососи) и некоторые ракообразные обладают обоими механизмами осморегуляции, могут жить как в пресной, так и в морской воде.
У пойкилосмотических животных (морские моллюски, иглокожие) осмотическое давление крови меняется параллельно с его изменением во внешней среде. Однако у этих животных имеется клеточная осморегуляция: при увеличении осмотического давления крови в клетках на такую же величину возрастает концентрация органических веществ (главным образом аминокислот), для которых клеточная мембрана слабо проницаема. В результате концентрация солей и содержание воды в клетке не меняются, осмотическое давление выравнивается за счёт накопления осмотически активных веществ. При уменьшении осмотического давления внешней среды и крови в клетках снижается концентрация органических веществ. Клеточная осморегуляция создаёт условия для ограниченной адаптации пойкилоосмотических животных к колебаниям осмотического давления во внешней среде.
Осморегуляция у пресноводных рыб. В организме пресноводных животных концентрация жидкостей внутренней среды обычно несколько ниже, чем у морских форм, но выше чем в окружающей воде. Пресноводные животные всегда гипертоничны по отношению к среде.
Благодаря этому вода осмотическим путем постоянно поступает внутрь организма. Концентрация солей в пресных водоемах колеблется в пределах 0-0,5%, в среднем составляя 0,2%. У некоторых пресноводных животных различные покровные образования затрудняют проникновение воды через кожу (панцири, чешуя). Однако полная изоляция организма от осмотического поступления воды невозможна.Осморегуляция организма пресноводных рыб сводится к тому, что вода поступает в организм осмотическим путем через жабры и слизистую пищеварительного тракта,а избыток ее выводится через почки. Активного питья воды не происходит. (Рис.48.)
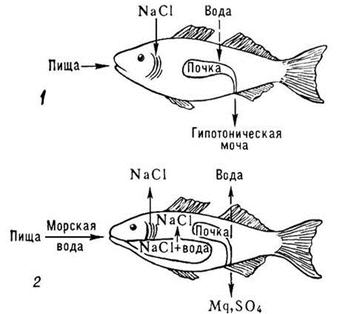
| Рис. 48. Осморегуляция у рыб: пресноводная рыба (1), морская костистая рыба (2); пунктиром обозначено движение воды по осмотическому градиенту. Пресноводные рыбы всасывают соли натрия жабрами; у морских костистых рыб клетки жаберного аппарата выделяют их. Почка морских рыб выделяет соли магния (сульфаты и др.), избыток которых поступает в организм рыб с пищей и морской водой. |
Таблица 15 Концентрация некоторых веществ в плазме крови водных позвоночных, ммоль/л.
| Виды | Среда обитания | Na | К | Мочевина | Осмотическая концентрация мосм/л |
| Морская вода Круглоротые Миога Mijxine Минога Petromyzon Минога Lampetra Хрящевые рыбы Cкат Raja Скат Potamotrygon Акула Sgualus Кистеперые рыбы Латимерия Latimeria Костистые рыбы Золотая рыбка Carassius Рыба-жаба Ospanus Угорь Anguilla >> >> Лосось Sabmo | море море пресные воды море пресные воды море море пресные воды море пресные воды море пресные воды море | - | - - | - - <1 <1 - |
Соли поступают с пищей и через жабры в процессе дыхания, а потеря их идет с мочой, экскрементами и частично через кожу. Фильтрационно-реабсорбционная функция почек, абсорбция, активность жаберного эпителия, зависит от соотношения разностей осмотических давлений воды и жидкостей организма.
Осморегуляция в море – у костных рыб. В результате морской трансгрессии многие материковые водоемы оказались связанными с океаном и осолонились. Условия водно-солевого обмена в морской и пресной воде кардинальным образом отличаются, в море организм животных оказывается гипотоничным по отношению к среде, что вызывает, постоянный отток воды из организма в окружающую среду и создает угрозу обезвоживания организма.
Приспособление к осморегуляции в таких условиях развивалось на основе пресноводного принципа строения почки, свойственного у большинства морских костных рыб. В этом случае функциональная активность клубочковой системы снижается, значительная часть клубочков вообще не участвует в фильтрации. Например, фильтрационная активность отдельных нефронов у форели в морской воде выше (3,74 нл/мин против 1,31 нл/мин в пресной воде), общая гломерулярная фильтрация за счет уменьшения числа функционирующих нефронов в морской воде ниже, чем в пресной (соответственно 20,1 и 142,6 нл/мин). У многих морских костистых рыб в соленой воде уменьшается размер клубочков, и их число. Среди морских костистых рыб есть антарктические виды, совершенно утратившие клубочки, а канальцы нефронов в таких агломерулярных почках заканчиваются слепо и обладают активной секреторной функцией. Утрата клубочков, способствует сохранению в крови гликопротеидов, играющих важную роль «биологических антифризов».
Снижение уровня почечной фильтрации не в состоянии компенсировать осмотические потери воды. Поэтому морские рыбы регулярно пьют воду, получая при этом избыточное количество солей. Угорь и морской подкаменщик поглощают за сутки до 50-200 см3 воды. Если блокировать ее поступления через кишечник, рыба обезвоживается и после потери 12-20% исходный массы погибает.
Реабсорбция ионов в почечных канальцах морских рыб снижена, зато здесь происходит обратное всасывание воды из канальцевой мочи. Соответственно морские рыбы выделяют сравнительно небольшое количество мочи, концентрация которой почти равна плазме крови. Интенсивно всасывается вода и в мочевом пузыре костистых рыб, стенки которого обладают высокой осмотической проницаемостью. Здесь абсорбируется хлористый натрий.
Избыток солей, кроме почек, выводится также через кишечник, здесь происходит интенсивное всасывание воды, а соли концентрируются и выводятся с фекалиями. Это относится главным образом к двухвалентным ионам, тогда как хлористый натрий активно абсорбируется, создавая в слизистой оболочке градиент концентрации, вследствие которого вода переносится через стенку кишечника.
Большую роль в выведении избытка солей играют жабры, одновалентные (главным образом Na+ и Cl-) экскретируются почти исключительно через жабры, выполняющие у рыб двойную функцию- дыхания и выделения. В жаберном эпителии есть особые крупные бокаловидные клетки. Эти «хлоридные» клетки расположены в первичных жаберных лепестках и в отличие от дыхательных клеток связаны с сосудами венозной системы. Перенос ионов через жаберный эпителий имеет характер активного транспорта и идет с затратой энергии. Стимулом экскреторной деятельности хлоридных клеток является повышение осмолярности крови.
Участие жабр в активном транспорте одновалентных ионов характерно не только для костистых рыб. Хлоридные клетки обнаружены у акуловых рыб, хрящевых и костных ганоидов, свойствен и пресноводным рыбам. Гипертоничность внешней среды обусловливает постоянные осмотические потери воды (в основном через жабры), которые восполняются путем питья. Избыток получаемых при этом солей выводится через почки и фекалиями (главным образом двухвалентные ионы), а также активно экскретируются особыми клетками жаберного эпителия (в основном Na+ и Cl-). Функционирование механизмов осморегуляции позволяет поддерживать осмотическое давление внутренней среды на относительно постоянном уровне, гипотическом по отношению к внешней среде.
Механизм осморегуляции у хрящевых рыб. Концентрация солей в их крови близка к таковой у костистых рыб и ниже, чем концентрация их в морской воде. Однако осмотическое давление жидкостей тела у этих рыб практически равно осмотическому давлению морской воды. В почечных канальцах хрящевых рыб активно реабсорбируется мочевина, 70-90% которой возвращается обратно в кровь, повышая ее суммарное осмотическое давление. Проницаемость жабр для мочевины понижена, и избыток мочевины выводится только через почки. Помимо мочевины, в крови хрящевых рыб накапливается значительное количество триметиламиноксида (ТМАО), обладающего высокой осмотической активностью. У костистых морских рыб содержание ТМАО в крови составляет 25-460мг %, то у акуловых оно достигает 250-1470 мг%.
На снижение солености среды хрящевые рыбы реагируют уменьшением реабсорбции мочевины и усилением выведения ее с мочой. Благодаря этим регуляторным процессам акуловые рыбы выдерживают больше колебания солености среды, появляясь даже в пресных водах.
Изоосмотичность морских пластиножаберных рыб не означает отсутствия ионной регуляции. Напротив, содержание солей в составе внутренней жидкой среды поддерживается на довольно постоянном уровне. Количество хлористого натрия в крови акул и скатов почти в два раза ниже, чем в морской воде, несмотря на постоянный приток этих ионов с пищей и благодаря диффузии через жабры. Часть избыточных солей выводится с мочой, концентрация ионов калия, кальция, магния и некоторых других в проксимальных сегментах нефронов нарастает.
Хлористый натрий, напротив, реабсорбируется в дистальных почечных сегментах нефронов и возвращается в кровь. Главную роль в экскреции хлористого натрия, видимо, играет ректальная железа – пальцеобразный вырост, соединенный протоком с клоакой. Показано, что до 75% выводимого из организма хлористого натрия экскретируется ректальной железой. Акуловые рыбы в отличие от костистых не пьют морскую воду.
Значительное количество мочевины синтезируется и задерживается в крови у двоякодышащих рыб. У осетровых рыб в морской воде также увеличивается содержание осмотически активных веществ сыворотки крови, что ведет к выравниванию осмотического давления крови и окружающей среды. При этом в отличие от акуловых рыб у ряда осетровых колеблется и содержание солей, в частности хлористого натрия. Осморегуляция у осетровых рыб происходит по промежуточному типу, между осморегуляцией хрящевых и костистых рыб.
Физиологический контроль осморегуляции. Механизмы регуляции водно – солевого обмена у рыб находятся в сложном взаимодействии. Водно – солевой обмен может лабильно изменяться в зависимости от динамики условий внешней среды и внутреннего состояния организма. Адаптивные возможности проявляются у разных видов неодинаково.
Примером широкой приспособляемости к солевому режиму водоемов служат так называемые проходные формы круглоротых (миноги) и рыб. При миграциях из моря в реки (или наоборот) механизмы осморегуляции у этих животных полярно преобразуются (смена гипо – и гипертонического состояния организма), что основано на пресноводном типе структуры почек у всех водных позвоночных.
У молоди лососевых рыб процесс подготовки к смене типа осморегуляции начинается еще в реке: увеличивается концентрация осмотических активных веществ, в плазме крови возрастает число хлоридных клеток в жабрах.
Таблица 16. Показатели осморегуляции у угря Anguilla amgnikka в реке и в море (по Строганову, 1962).
| Водоем | D to среды | Выделение мочи (мл\кг в сутки). | D to мочи | Dto крови |
| Река | 0,08 | 60-150 | 0,09 | 0,63 |
| Море | 1,85, | 2-4 | 0,79 | 0,82 |
Все это повышает устойчивость к возрастающей солености воды при выходе в море. У взрослых особей перестройки носят обратный характер, что позволяет им подняться вверх по течению на месте нереста. Все эти изменения стимулируются нейроэндокринными механизмами.
А ктивная осморегуляция обеспечивает не только приспособления принципиального характера (регуляция в пресной или морской воде), но и лабильные адаптивные реакции на изменение градиента плотности. У рыб регуляция этих процессов связана главным образом с нервно-гуморальными механизмами системы гипоталамус – гипофиз – интерреналовая ткань.
Гипоталамус представляет центральную структуру головного мозга, в которой сигналы с осморецепторов трансформируются в реакции организма. Вырабатываемые в ответ на осмотическую стимуляцию нейросекреты гипоталамуса переносятся по аксонам в заднюю долю гипофиза (нейрогипофиз), откуда в виде гормонов поступают в кровь. У рыб в нейрогипофизе продуцируются гормоны аргинин – вазотоцин (АВТ), и изотоцин (ИТ), окситоцин (ОТ) и вазопрессин (ВП). Первые три гормона в низких дозах (до 1 мг/кг) действуют в пресной воде как антидиуретики, а в морской воде их влияние на диурез не проявляется. В высоких дозах (более 10 мг/кг) гормоны повышают диурез.
Гормон вазопрессин, обнаруженный у рыб, обладает диуретическим действием, причем в пресной воде он подавляет антидиуретический эффект гормонов аргинин – вазотоцина (АВТ), изотоцина (ИТ) и окситоцина (ОТ2). Все нейрогипофизарные гормоны влияют на диурез, воздействуя на интенсивность клубочковой фильтрации.
Другая форма влияния гипоталамуса на водно-солевой обмен заключается в стимуляции эндокринной деятельности передней доли гипофиза (аденогипофиз). Проникающие в аденогипофиз нейросекреты стимулируют определенные группы клеток, продуцирующих гормоны.
Наибольшее заключение в осморегуляции имеют пролактин и адренокортикотропный гормон (АКТГ). Пролактин играет роль в пресноводной осморегуляции, секретируется большем количестве у пресноводных рыб, чем у морских. Стимуляция других клеток аденогипофиза влечет за собой увеличение секреции АКТГ, которая в свою очередь повышает активность интерреналовой ткани, клетки которой выделяют ряд кортикостероидов, принимающих непосредственное участие в регуляции водно-солевого обмена. В интерреналовой ткани костистых рыб вырабатываются не менее шести кортикостероидов, а также ряд их производных. Наиболее значительную роль в регуляции водно-солевого обмена играет гормон кортизол, в меньшей степени - альдостерон. (Рис. 49).
Кортизол восстанавливает осморегуляторные свойства и считается главным фактором приспособления рыб к осморегуляции в морской воде. У акулообразных и осетровых рыб гормональный контроль осморегуляторных процессов осуществляется аналогично.
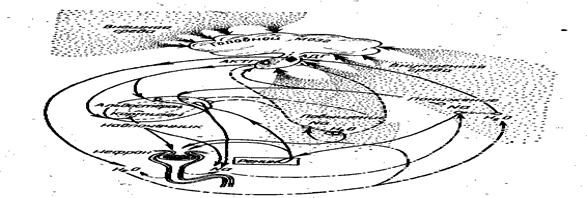
Рис. 49. Механизм гуморальной регуляции водно – солевого баланса
6.9 Обмен веществ в целом Общий обмен веществ не является постоянным и неизменным. Он значительно изменяется в зависимости от состояния организма и внешних условий.
На обмен веществ могут оказать влияние условия среды обитания рыб и характер питания, температурный режим воды и такие факторы, как возраст, рост, вес тела и пол. Функциональное состояние отдельных систем и органов также сказывается на интенсивности обмена веществ, особенно при заболевании. Ряд внешних условий, например, температура, барометрическое давление, состав и влажность воздуха, солнечная энергия и пр., может также резко влиять на обмен веществ.
О величине процессов обмена можно судить по продуктам распада, выделяемым из организма. Учёт азота в моче позволяет выяснить количество распавшегося в организме белка. Представление о распаде жиров и углеводов можно составить по выделению из организма углерода в форме СО2, так как конечным продуктом распада этих веществ является углекислый газ. Выделение СО2 из организма происходит через жабры и полужабры. Для учета выделенного СО2 и поглощённого О2 в физиологических экспериментах животных помещают в респирацион-
ную камеру. Зная количество выделенного азота и СО и поглощённого кислорода, можно составить представление об интенсивности обмена и степени использования организмом питательных веществ.
Дыхательный коэфициент — это соотношение между объёмами выделенного С02 и поглощённого О2. Он бывает различным: при окислении углеводов он равен 1, при распаде жиров — 0,7, а при распаде белков — 0,8.
Обмен веществ при голодании. При голоде организм не получает извне питательных веществ: Поддержание основных процессов, обеспечивающих жизнь организма, происходит за счёт распада запасов питательных веществ. Первый период голодания характеризуется распадом веществ, полученных организмом с воспринятой до голодания пищей.
С первых дней голодания распад белков значительно снижается, а в последующие дни расщепление белка поддерживается на низком уровне. Энергетические затраты происходят за счёт гликогена, который защищает белок от гибели. Когда запасы гликогена потрачены, организм живёт за счёт своего жира. Этот период зависит от богатства жиром данного животного. Поэтому жирные организмы голод переносят дольше, чем худые. При исчезновении запасов жира, как источника энергии, наблюдается повышенный распад тканевого белка, который организм оберегал от распада. Этот период усиленного распада тканевого белка является предвестником голодной смерти.
Во время голодания органы уменьшаются в своей массе, но не в равной степени. Больше всего исчезает жировая ткань (до 93—97%), затем следуют мышцы и железы (40—70%). Меньше теряет в весе центральная нервная система (2—3%). Таким образом, происходит сохранение органов, более важных для жизни. Смерть наступает, по некоторым данным, после потери животным 20—50% своего веса.
Обмен при голодании даёт представление о способности организма к ограничению расходования белков, как основного структурного материала клеток, а также о компенсированном и последовательном распаде запасов углеводов и жиров. Такой обмен служит показателем минимального расходования веществ, необходимых для пополнения затрат на основные процессы, поддерживающие жизнь (работа сердца, дыхательных мышц, печени, нервной системы), и на регуляцию тепла.
БЕЛКОВЫЙ МИНИМУМ. Белок пищи постоянно идёт на восстановление тканевого белка и на образование регулятора жизненных процессов, например, гормонов. В питании белок нельзя заменить ни углеводами, ни жирами, поскольку они не содержат азота. Если количество белка в пище будет убавлено ниже необходимого минимума, организм уже не сумеет сохранить азотистое равновесие, и начнёт тратить свои тканевые белки. Баланс азота в этом случае становится отрицательным.
Белковый минимум, это минимальное количество белка, которое является необходимым для существования организма. Количество белка в пище должно быть выше минимальной потребности организма в азотистых соединениях. Организм всегда должен иметь некоторый резерв, который можно использовать в случае усиления физиологической деятельности.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 3442; Нарушение авторских прав?; Мы поможем в написании вашей работы!