КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физиология органов размножения рыб
|
|
|
|
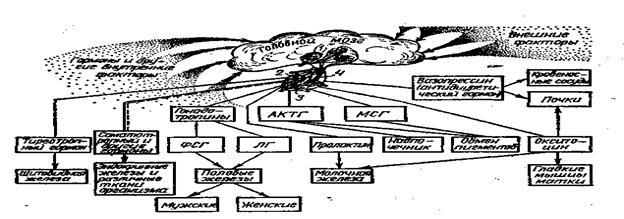
Ядра гипоталямуса; 2-передняя доля гипофиза;3-средняя доля гипофиза;4-задняя доля гипофиза.
 8.2Щитовидная железа. Тироксин и трийодтиронин – гормоны щитовидной железы. Известное свойство тирозина – его способность присоединять йод, участвующий в регуляции обмена веществ
8.2Щитовидная железа. Тироксин и трийодтиронин – гормоны щитовидной железы. Известное свойство тирозина – его способность присоединять йод, участвующий в регуляции обмена веществ
Клетки щитовидной железы, производящие тироксин и трийодтиронин - это тирроидные фолликулы, рассеяные вдоль спинной аорты, в тканях селезенки, стенках спинной аорты. Данные фолликулы выделяют три гормона; -Т2;-Т3;-Т4. Они происходят из другого зачатка, чем те, которые вырабатывают кальцитонин. Консолидация однородных элементов в единый орган и ассоциация разнородных желез в одну, служит целям лучшей регуляции их деятельности. Паращитовидная железа появилась на уровне амфибии, у рыб она отсутствует.
Паращитовидная железа появилась только на уровне амфибии,у рыб она отсутствует
8.3Поджелудочная железа. Рассеянные вдоль кишечника эндокринные клетки занимают большую поверхность и дальнейшее разрастание их шло бы в ущерб поверхности всасывания. Поджелудочная железа, выполняет двойную функцию – секреторную и инкреторную (смешанную). Инкреторная функция заключается в выработке островками Лангерганса гормона инсулина и его антогониста глюкагона. Гормон инсулин регулирует углеводно – жировой обмен, понижая уровень сахара в крови, а глюкагон выполняет противоположную функцию.
У костистых рыб она обьединена с печенью и носит название – гепатопанкриеса, у хрящевых рыб представлен, как компактный орган. Клетки эндокринной системырыб объединяются с поджелудочной железой или образуют отдельные железки, состоящие из островковой ткани. Примером ассоциации является объединение хромаффинной и интерренальной тканей в одну железу - в надпочечник. У хрящевых рыб оба органа еще разъединены. Интерренальная железа расположена между почками, почему и получила свое название. У костистых рыб оба органа уже расположены в области кардинальных вен и соприкасаются друг с другом.
8.4Интерреналовые тельца являются аналогами корковой части надпочечников у высших животных. Морфологически они вкраплены в ткани головной почки. В интерреналовой ткани костистых рыб вырабатывается не менее шести кортикостороидов, а также ряд их производных. Наиболее важное значение в регуляции водно-солевого обмена играет кортизол и кортикостерон, в меньшей степени – альдостерон. Кортизол – выполняет стрессоподавляющую функцию, усиливает распад и накопление гликогена в печени, поддерживая постоянство концентрации глюкозы в крови. У пресноводных рыб данный гормон регулирует водно – солевой баланс, восстанавливая осморегуляторные свойства, который считается главным фактором приспособления рыб к осморегуляции в морской воде. У акулообразных и осетровых рыб гормональный контроль осморегуляторных процессов осуществляется аналогично. Кортикостерон участвует в осморегуляции, регулирует работу почек
8.5 Гормоны надпочечников Альдестерон у круглоротых по – видимому, отсутствует, обнаружен лишь у некоторых рыб, в том числе у южноамериканской двоякодышащей рыбы. Деление кортикоидов на глюкокортикоиды и минералокортикоиды правомерно только для высших позвоночных. У низших позвоночных, глюкокортикоиды выполняют также и минералокортикоидную - смешанную функцию.
Определенную роль в регуляции водно-солевого обмена у рыб играют гормоны, выделяемые клетками хромаффиновой ткани, особенно адреналин (эпинефрин). Адреналин увеличивает проницаемость жаберного эпителия для воды, одновременно снижается выведение хлоридов и натрия через жабры, исполняет роль стресс гормона, выбрасываясь в кровь во время сильного стресса.
У морских рыб кортикоиды стимулируют функцию «хлоридных» клеток, удаляющих соль из организма. У пресноводных рыб, которые извлекают соль из наружной среды, они способствуют этому процессу и вместе с тем помогают задерживать натрий в организме. В почках они способствуют реабсорбции натрия.
8.6Гормоны гипофиза. Гипофиз, участок головного мозга, выполняет функцию желез внутренней секреции, является мотором всей эндокринной системы в целом. Морфологически гипофиз делится на три отдела у высших позвоночных животных, а у рыб различают два отдела, среднего отдела у рыб нет.
У позвоночных нейрогормоны выделяются преимущетсвенно в области терминалей дендритов и аксонов гипоталямо-гипофизарной системы. Поступая в кровь, спинномозговую и тканевую жидкость. Посредством этих жидкостей нейрогормон достигают клеток- мишеней и оказывают на них регулирующее влияние это аргинин – вазотоцин (АВТ), изотоцин (ИТ) и окситоцин (ОТ-2).
Все нейрогипофизарные гормоны влияют на диурез, воздействуя на интенсивность клубочковой фильтрации. Другая форма влияния гипоталамуса на водно - солевой обмен заключается в стимуляции эндокринной деятельности передней доли гипофиза (аденогипофиз). Нейросекреты, проникающие в аденогипофиз стимулируют определенные группы клеток, продуцирующих гормоны. Наибольшее значение в осморегуляции имеют пролактин и адренокортикотропный гормон (АКТГ). Адренокортикотропный гормон также регулирует процессы адаптации к стрессам, являясь антагонистом к действию адреналина. У некоторых рыб гормон пролактин вызывает утолщение эпителия и выделение кожной слизи. Кроме того, гормон поддерживает инстинкт заботы о потомстве, даже у живородящих рыб. Пролактин имеет значение для поддержания водно- солевого равновесия. Удаление гипофиза эвригалинных карпозубых рыб делает их неспособными жить в пресной воде из-за чрезмерной потери ионов натрия жабрами и через кожу, что приводит к падению осмотического давления крови. Введение пролактина спасает рыб от гибели. Пролактин играет ведущую роль в пресноводной осморегуляции. Гормон пролактин секретируется у пресноводных рыб в большем количестве, чем у морских.
Задней долей гипофиза (нейрогипофиз), вырабатывается гормон- окситоцин (ихтиотоцин), обладающим осморегуляторной и свойствами регуляции образования слизи. Вазопрессин – вазотоцин, регулирующий работу сердечно- сосудистой системы, тонус кровеносных сосудов и водно - солевой обмен. Меланофорный гормон (МФГ) - гормон регулирующий пигментацию, обычно обеспечивает темную окраску кожи.
Соматотропный гормон (СТГ) – регулирует рост и развитие организма, регенерацию тканей и органов.
Тиреотропный гормон (ТТГ) – регулирует работу щитовидной железы
Стимуляция других клеток аденогипофиза влечет за собой увеличение секреции АКТГ, в свою очередь повышающего активность интерреналовой ткани, клетки которой выделяют ряд кортикостероидов, принимающих непосредственное участие в регуляции водно – солевого обмена.
Помимо интерреналовой и хромаффиновой ткани, в почках лучеперых рыб обнаруживаются отдельные группы железистых клеток, заключенных в соединительнотканные капсулы – тельца Станниуса. Эти образования участвуют в регуляции обмена кальция и водно – солевого обмена, вырабатывают вещество гипокальцин.
Для рыб характерна еще каудальная нейросекреторная система: она находится в заднем отделе спинного мозга, где расположены нейросекреторные клетки, отростки которых оканчиваются в его брюшной части. Вырабатывает гормон ренин, регулирующий кровянное давление – ангиотензин 2.
Участок спинного мозга, в котором происходит накопления секрета, и последующее выведение его в кровь носит название урофиза (или нейрофиза). У костистых рыб урофиз вычленяется в виде вентрального выроста спинного мозга. У хрящевых и осетровых рыб он морфологически не выражен, однако система нейросекреторных клеток имеется. Структурная организация урофиза и нейрогипофиза очень сходны между собой. Есть сведения и об участии этой системы в регуляции водно - солевого обмена: введение экстрактов урофиза карпа лабораторным крысам продемонстрировало антидиуретический эффект. (Zshikame, 1970). Показано, что урофиз влияет на уровень ионов натрия, магния и хлора в плазме крови (Fryer et. al., 1978). Вещества, выделяемые урофизом называют – уротензинами. Уротензины регулируют давление крови, сокращение гладкой мускулатуры.
8.7 Половые гормоны. Половые гормоны выполняют смешанную секрецию, ястыки самок рыб вырабатывают в больших количествах гормоны эстрогены определяющих пол, половое поведение, развивают вторичные половые признаки у рыб.
Семенники самцов вырабатывают мужские половые гормоны – тестостерон, стимулирующий выработку и секрецию спермии формирующие сперматогенез, половое поведение и вторичные половые признаки.
Контрольные вопросы.
1.Обьясните понятие гормоны и инкреты.
2.Как квалифицируются гормоны по классам и функциональным признакам?
3.Поясните механизм функциональной взаимосвязи эндокринных желез.
4.Роль щитовидной железы и их гормонов.
5.Какие функциональные изменения наступают в организме рыб при дефиците или избытке инсулина?
6.Какова роль гипофиза в эндокринной системе?
7.Принципы практического использования гормонов в рыбоводстве.
Глава 9
9.1 Размножение рыб. Сохранение и увеличение рыбных запасов в естественных водоемах направлены в основном на обеспечение размножения рыб.
Размножение – это звено жизненного цикла рыбы, обеспечивающее во взаимодействии с другими звеньями воспроизводство популяции и сохранение вида. Для каждого вида рыб характерны свои специфические особенности размножения. Они обусловлены адаптацией разных видов рыб к определенным условиям размножения и развития молоди, которая дает пополнение, необходимое для сохранения вида и поддержания его численности.
У одного и того же вида рыбы плодовитость подвержена весьма большим изменениям и, в первую очередь, в связи с изменением размеров рыбы. У большинства рыб количество икринок с возрастом сначала увеличивается, а затем, с началом старения особи, уменьшается. Сильные изменения претерпевают плодовитость у одной и той же популяции в зависимости от обеспеченности пищей.
Для рыб с порционным типом икрометания, характерно одновременное созревание лишь части икры в яичнике. Промежутки между выметыванием отдельных порций икры, часто связано с продолжительностью первого этапа внешнего питания личинки. Когда личинка первой порции икрометания достигает второго этапа внешнего питания и переходит на иной вид корма, личинки следующей порции нереста только начинают внешнее питание. У большинства рыб наиболее высококачественное потомство получается от рыб среднего возраста. Молодые и очень старые дают менее жизнестойкое потомство. Первые порции спермы у самцов более высококачественные. Среди рыб одного поколения быстрее созревают особи с большей жирностью тела, однако рано созревающие рыбы обладают меньшей воспроизводительной способностью, чем поздно созревающие: плодовитость их ниже, икринки мельче, с меньшим количеством жира.
9.2 Жизненный цикл рыб. Жизненный цикл рыб распадается на различные периоды, каждый из которых характеризуется определенными морфологическими и физиологическими особенностями.
Эмбриональный период - эндогенное питание, длится от момента оплодотворения до перехода на экзогенное питание. Подразделяется на два подпериода:
- собственно эмбриональный, когда развитие происходит внутри яйцевой оболочки,
- предличиночный, когда выклюнувшиеся свободноживущие особи питаются за счет желтка.
Личиночный период - переход на активное внешнее питание с сохранением личиночных органов и признаков.
Мальковый период - по внешнему строению организм преобретает сходство со взрослым. Закладывается чешуя. Дифференциация пола, но половые органы не развиты.
Ювенильный период - усиленное развитие половых желез, но рыбы еще неполовозрелы. Начинают развиваться вторичное половые признаки.
Взрослый организм - с момента половой зрелости, делится на с тарческий период - замедляется или полностью прекращается рост. Рыба теряет способность размножаться.
Эмбриональный период в свою очередь разделяется на несколько стадий:
I «а» стадия - начало дробления. I стадия - дальнейшее дробление, гаструляция (обрастание желтка зародышевым слоем), до появления зародышевой полоски.
II стадия - образование зародышевой полоски. Дифференцировка туловищных миотомов. Стадия заканчивается отделением хвостовой почки от желтка.
III стадия - несформировавшегося эмбриона. Начинается от момента обособления хвоста и длится до охвата эмбрионом всего желтка.
IV стадия - сформировавшегося эмбриона. Начинается пульсация сердца и подергивание эмбриона. Длится до выхода эмбриона из икринки.
9.3 Половая система рыб. Рыбы, как правило, раздельнополы. Однако некоторые рыбы двуполы. Бывают случаи превращения одного пола в другой. Половые железы самца представлены парными семенниками (молоками), содержащими в период размножения огромное количество сперматозоидов. От семенников идут протоки, которые открываются в половое отверстие.
Половые железы самки состоят из парных (как исключение, из одного) яичников, переходящих в короткие выносящие протоки, открывающиеся в половое отверстие. В яичниках находятся икринки (яйца). Количество икринок зависит как от возраста и размеров рыб (чем старше и крупнее рыба, тем больше икры), так и от их биологических особенностей (как правило, чем меньше данный вид рыбы проявляет заботы о потомстве, тем больше икры мечут самки).
Икринки в большинстве своем очень мелкие, круглые, богатые желтком. Строение икринки (яйцеклетки) костистых рыб довольно сложное. Икринка покрыта двумя оболочками: внешняя часть бывает студенистой и липкой; внутренняя толстая оболочка иногда состоит из двух слоев – более плотного и менее плотного. Оболочки имеют отверстие – микропиле, сквозь которое внутрь икринки проникают сперматозоиды. Помимо двух оболочек имеется зародышевый диск, состоящий из протоплазмы с ядром. Запас белкового вещества сосредоточен в желтке, в определенном месте которого находится одна или много жировых капель.
Оплодотворение у подавляющего большинства рыб происходит вне тела, в воде. Процесс оплодотворения заключается в том, что сперматозоиды проникают внутрь икринки через микропиле, ядро яйцеклетки сливается с ядром сперматозоида и начинается дробление, ведущее к развитию зародыша. Зародыш развивается на питательном желтке, часть желтка остается в качестве резервного материала в виде желточного мешка у личинки.
Рыбы, как правило, раздельнополы. У костистой рыбы в верхней части брюшной полости по бокам плавательного пузыря располагаются парные половые железы. У самок половые железы представлены длинными яичниками (ovaruem), которые имеют хорошо заметную «зернистую» структуру. Задние, вытянутые отделы яичников играют роль выводных протоков и открываются непарным половым отверстием позади анального отверстия.
Половые железы самцов – длинные, гладкие, довольно плотные семенники (testis). Задние отделы семенников превратились в короткие выносящие протоки, открывающиеся общим половым отверстием позади
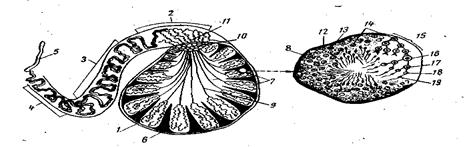
Рис.58. Схема строения семеника и поперечного разреза извитого канальца:
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 2352; Нарушение авторских прав?; Мы поможем в написании вашей работы!