КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Некоторых видов рыб
|
|
|
|
| Вид рыбы | Длина, см. | Максимальная, известная частота, Гц | Вид рыбы | Длина, см. | Максимальная известная частота, Гц | |
| Тунец Вьюн Бычок Карп | Хамса Атерина Колюшка Гольян | |||||
У крупных хищников и мигрантов абсолютные значения максимальной скорости могут достигать больших величин. Двухметровая рыба, работающая с частотой 30 Гц и движущаяся со скоростью около 20 длин тела в секунду, развивает скорость 40 м/с, или 144 км/ч. Одиночные мощные разгибы тела могут происходить в течение промежутков времени короче 1/30 сек. В этот момент может происходить еще более быстрое — до 40 длин тела в секунду, но очень кратковременное перемещение. По аналогии со спортом такие рывки можно назвать прыжками с разбега.
10.4 Полет рыб. Среди рыб есть представители, которые приспособились к своеобразному полету по воздуху, это не настоящий полет, а парение по типу планера. У этих рыб грудные плавники чрезвычайно сильно развиты и выполняют ту же функцию, что крылья самолета или планера. Основной двигатель, дающий начальную скорость при полете, хвост и в первую очередь его нижняя лопасть. Выскочив на поверхность воды, летучая рыба еще некоторое время скользит по водной поверхности, оставляя за собой кольцевые волны, расходящиеся в стороны. В то время, когда тело летучей рыбы находится в воздухе, а в воде остается только ее хвост, она все еще продолжает увеличивать скорость движения, нарастание которой прекращается только после полного отрыва тела от поверхности воды.
Держаться в воздухе летучая рыба может около 10-15 сек. поднимаясь в высоту до 4м. над поверхностью воды, пролетая при этом расстояние свыше 100 м. Дальше держаться в воздухе могут те рыбы, у которых увеличены как грудные, так и брюшные плавники.
В момент взлета грудные плавники рыбы широко раскрыты и расположены под углами к направлению движения, благодаря чему создают необходимую подъемную силу. Эта функция и обусловливает их очень сильное развитие.
Полет у летучих рыб выработался как защитное приспособление, позволяющее им ускользать от хищников - тунцов, корифен, меч - рыб. Полет рыб есть так же приспособление к перемещению с меньшей затратой энергии от одного скопления кормовых планктонных организмов к другому.
Среди харациновых рыб есть представители, приспособившиеся к активному машущему полету. Это мелкие рыбки до 9-10см длины, населяющие пресные воды южной Америки. Они могут выскакивать из воды и пролетать при помощи взмахов удленненных грудных плавников до 3-5 метров. Хотя у летающих харацинид размеры грудных плавников меньше, чем у летучих рыб данного семейства. Грудные мышцы, приводя в движение грудные плавники, хорошо развитые у харациновых рыб, приспособившихся к очень сильно развитым костям плечевого пояса, которые образуют некоторое подобие грудного киля птиц. У летающих рыб плавники достигают 25% веса тела, в то время как у нелетающих представителей близкого рода - только 0,7%.
Другим способам движения рыб можно отнести движения при помощи прикрепления к плавающим предметам паразитизма, комменсализма и симбиоза. Изменением направления двигателя, сохранением равновесия и торможением, морфологически касаются главным образом особенностей строения плавников. Обособляются плавники, выполняющие функции несущих плоскостей, рулей килей стабилизаторов и локомоторных органов, причем расположение всех этих плавников на теле строго подчинено генетическому коду.
10.5 Общая характеристика скелетной мускулатуры.
Движение—одно из проявлений жизни; у позвоночных животных оно осуществляется специальными органами —мускулами, построенными из поперечнополосатой мышечной ткани. Мускулы закрепляются на скелете на системе рычагов и в целом составляют скелетную мускулатуру.
Своей работой скелетная мускулатура обеспечивает: а) перемещение тела животного в пространстве (в поисках пищи, особи другого пола, в целях защиты); 6) изменение положения отдельных частей тела относительно друг друга; в) изменение объема полостей тела (при дыхании и пр.); г) изменение положения и направления складок кожи (движение рта и т. д.).Движения животного являются ответом на раздражения, полученные из внешней среды, мышечные движения — одно из внешних проявлений мозговой деятельности (И. М. Сеченов).
Мышечная система в значительной степени влияет на строение и развитие органов активного движения в целом, и в зависимости от степени и характера ее собственного развития происходят соответствующие изменения в строении скелета.
Наконец, мышечная система служит трансформатором химической энергии, поступающей в организм в виде пищи, в тепловую (до 70%) и в механическую работу (всего лишь около 30%).
10.5.1 Морфология и физиология мышц (мускула). М ускул, или мышца,—musculus—имеет соединительнотканный остов и главные функционирующие элементы, которыми служат поперечнополосатые мускульные волокна.
Соединительнотканному остову присущи общие для всех органов черты построения, т. е. снаружи мускул одет соединительнотканной оболочкой—наружным перимизием—perimysium extcrnum s. epimysmm — который иногда содержит небольшое количество жировой ткани. От этой оболочки проходят внутрь органа значительные по толщине перегородки, или; трабекулы (2), связывающие между собой крупные мускульные пучки. Перегородки, в свою очередь, отдают от себя ряд более тонких перегородок, которыми охватываются менее крупные мускульные пучки. Все эти внутренние переплеты, по которым ветвятся сосуды и нервы, получают название внутреннего перимизия—perimysium internum (3). Между мускульными волокнами самых мелких мускульных пучочков также пробегают очень нежные прослойки, называемые ондомизием—endomysium. Оболочка вместе со всеми указанными перегородками и составляет остов мускула, в который закономерно вплетены мускульные волокна. Все эти образования в совокупности и дают различной величины и формы мускульное брюшко.
Каждое мускульное брюшко имеет сухожильный добавок—сухожилие. Форма сухожилия зависит от формы мускульного брюшка. Если мускульное брюшко в сечении округлой формы, то и сухожилие как его добавок становится шнурообразным. В таких случаях его принято называть просто сухожилием- tendo (tenon). Если же мускул плоский, то он переходит в пластинчатое сухожилие-апоневроз.
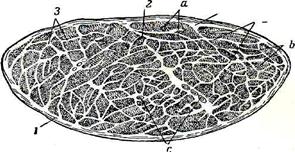
| Рис.69. Поперечный разрез мускула. 1— оболочка мускула, или наружный перимизий; 2 —трабекулы,3- внутрен -ний перимизий, разграничивающий крупные мускульные пуши - пучки (а); Л—внутренний перимизий, разгра- ничивающий менее крупные пучки (Ь); с—сосуды и нервы. |
Сухожильные добавки построены наподобие самого мускульного брюшка, но вместо пучков сократительных элементов здесь находятся
параллельные волокна плотной оформленной соединительной ткани. Эти волокна идут пучками и заложены в свой остов, который является продолжением остова брюшка, т. е. оболочка и трабекулы брюшка—наружный и внутренний перимизии—переходят в оболочку и трабекулы сухожилия— peritendmeum (peri-tenonium) extcrmim et peritendmeum internum. Волокна сухожилия крепко соединены с мускульными волокнами (а может быть, и переходят непосредственно друг в друга). Сухожилие становится очень плотным (компактным) и прочным шнуром благодаря наличию лишь крайне незначительной соединительнотканной прослойки между строго параллельно направленными коллагенными пучками волокон, скудно снабженными сосудами. Все это делает сухожилие мало утомляемой частью мускула. Кроме того, сухожилие по сравнению с мускульным брюшком обладает огромной сопротивляемостью на растяжение (Рис.70).
Сухожилия весьма крепко соединены с рычагами движения, проникая в виде шарпеевских волокон в толщу костного вещества, с такой конструкцией и свойствами сухожилие принимает на себя функцию передатчика силы сокращения мускульного брюшка на отдаленные, скелетные части, т. е. играет роль трансмиссионного приспособления.
Вступающие в мускул и выходящие из него сосуды распределятся в глубину по перекладинам остова. Артерии, подходя к мускульным волокнам, образуют густую сеть капилляров, петли которой растянуты в направлении длинной оси волокон.
Мускул снабжен двигательными и чувствительными нервными волокнами. Каждое отдельное волокно мускула управляется только ему присущим самостоятельным двигательным волокном и вместе с ним представляет одну целую, очень маленькую рабочую единицу. Это единство устанавливается у зародыша с самых начальных стадий развития мускульных клеток (волокон).
Таким образом, комплекс объединенных мускульных волокон дает вместе с остовом и сухожилием орган-мускул, который, согласно сказанному, неразрывно связан со своими нервными волокнами, составляющими целый нерв данного мускула.
В процессе филогенеза при функциональной дифференцировке мускула последний может распадаться на новые мускульные единицы, и этому распадению обязательно следует и сопровождающий его нерв. При обратных преобразованиях, т. е. при слиянии в результате объединения работы нескольких мускулов в один сплошной орган, к нему будут подходить, и его будут иннервировать несколько нервов, и мускул станет по происхождению сложным. Если принять в соображение, что каждый мускул развивается из определенного мускульного сегмента (миотома или его вентрального отпрыска) и иннервируется нервом того же сегмента, то легко понять, что иннервация мускулатуры различными ветвями служит очень ценным критерием при определении происхождения и гомологии отдельных мускульных единиц.
Форма мускулов. Величина и форма мускулов чрезвычайно разнообразны и тесно связаны с взаиморасположением плечей рычагов, на которых укрепляются мускулы, и силой сокращения в том или другом месте организма.
Мускулы особенно рельефно обрисованы на свободных конечностях. По сравнению с мускулами туловища и головы они имеют в большинстве случаев удлиненную, в сечении округлую форму, напоминая или цилиндр, или конус, или веретено. На поясах конечностей эта рельефность формы и обособленности уже несколько стушевывается, а мускулы, простирающиеся между туловищем и плечевым поясом, характеризуются в большинстве пластинчатой формой, т. е. представляют так называемые широкие мускулы. Мускулы, располагающиеся вокруг отверстий, бывают кольцеобразной формы(Рис.71).
Внутреннее строение мускулов. Внутреннее строение мускульных брюшков бывает довольно разнообразно, наиболее простым видом строения следует считать брюшко, у которого направление волокон параллельно длинной оси мускула. Такого типа строения мускулы очень мясистые и называются по функции динамическими, а по строению простыми. Мускул тем сильнее, чем больше в нем мышечных волокон. Количество волокон практически сосчитать нельзя, поэтому силу мускула принято определять путем измерения его физиологического поперечника. В простых мускулах, как уже отмечалось, все пучки мышечных волокон параллельны длинной оси мускула; следовательно, в данном случае физиологический поперечник совпадает с физиологическим и анатомическим поперечником, который также перпендикулярен длинной оси мускула.
В зависимости от гистологического строения подъемная сила мускула может колебаться в пределах от 5 до 15 кг (в среднем 8—10 кг) на 1 см² физиологического поперечника. Расстояние, на которое мускул может проявить свою силу, зависит от длины мышечных волокон, которая не превышает 12—15 см. Так как мышечное волокно при своем сокращении укорачивается приблизительно на половину своей длины, то мускул, имеющий физиологический поперечник в 1 см2, способен поднять груз в 10 кг на высоту 6—7 см, т. е. механическая работа мускула в этих условиях будет равняться приблизительно 0,6 кг/м.
Мускулы описанного динамического типа, будучи маломощными, встречаются крайне редко. Чаще мышечные волокна имеют косое направление по отношению к длинникумускула, располагаясь при этом в нескольких вариациях.
1. Начальное и концевое сухожилиямогут покрывать мускульное брюшко с его противоположных поверхностей на большем или меньшем протяжении. Они обычно хорошо видны и за свой блестящий белый цвет называются мышечными (сухожильными) зеркалами. В отношении к длиннику мускула пучки мышечных волокон идут косо от одного сухожилия к другому или от кости (если мускул начинается мясисто) к концевому сухожилию. Такие мускулы называются одно-или полуперистыми— m. unipermatus. Вполне понятно: а) что при одинаковых объемах простого и перистого мускулов количество мышечных волокон в последнем будет больше, но длина соответственно меньше. Следовательно, мускул будет выигрыватьв силе, но проигрывать в расстоянии; б) что физиологический поперечник (с~с) в перистых мускулах не будет соответствовать анатомическому (в — в) — последний меньше.
2. Одно из сухожилий может расщепляться на две пластины, которые покрывают один конец мышечного брюшка, а в толщу последнего внедряется противоположное сухожилие. В таком случае мышечные волокна будут косо подходить к центральному сухожилию с двух противоположных поверхностей. Такой мускул по своему строению—д вуперисты й—m. bipennatus. В нем будет еще больше мышечных волокон, а длина, напротив, еще меньше. Двуперистый мускул еще больше выигрывает в силе за счет большого проигрыша в расстоянии. Наиболее сложное устройство приобретает мускул, когда расщепляет сухожилия; например, одно сухожилие на три пластины, и тут создается громадная подъемная сила, действующая, на очень коротком расстоянии.
Двухперистые и многоперистые мускулы называются статодинамическим и, так как они по преимуществу выполняют статическую функцию (например, при упоре животного). Вместе с тем такие мускулы (перистые) при соответствующих условиях могут давать также очень значительный размах. К таким условиям может быть отнесено использование более короткого плеча силы рычага, чем это требуется для динамического мускула. При этом такие мускулы дают очень сильный, хотя и короткий первоначальный импульс движению, который обычно совпадает с моментом наибольшей потребности в силе сокращения и способен по инерции увеличить размах плеча сопротивления.
Рис. 70. Типы построения мускульных брюшков
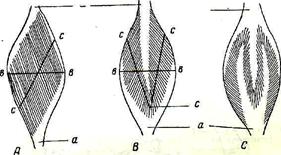
А - одноперистый тип;
Б - двуперистый тип;
С - многоперистый тип;
а- сухожилие;е-е- анатомический
и с- с- физиологический поперечник.
Механизм действия мускулов. Действие мускулов выражается их сокращением, причем химическая энергия мускульной тканью превращается в работу с выделением теплоты. При сокращении брюшко мускула укорачивается по длинной оси приблизительно наполовину, а в перистых мускулах очень мало, и утолщается в поперечнике. Это влечет за собой сближение пунктов прикрепления на рычагах, а следовательно, и самих рычагов (рис.70). Такое сближение достигается различными способами.
Во-первых, как это бывает чаще всего, один из пунктов прикрепления остается неподвижны м-punctum fixum-и в результате сокращения перемещается лишь та кость или часть тела, на которой находится подвижное прикрепление-punctum mobile.
Во-вторых, наблюдаются случаи, когда места прикрепления в функциональном отношении могут изменяться, т. е. неподвижное место при одной потребности передвижения может стать подвижным, а при другой потребности, наоборот, неподвижным.
Существуют и такие условия, при которых начало и конец мускула совсем отсутствуют, как, например, у мускулов, окружающих кольцом отверстия (ротовое) или полые органы (сфинктер прямой кишки у анального отверстия).
Закономерные действия мускулов или их групп в системе органов произвольного движения обусловливают самые разнообразные изменения в соотношении частей тела друг с другом и всего организма с внешней средой (перемещение по суше, в воде, в воздухе).
В частности, мускульная работа вызывает движения: 1) друг относительно друга частей позвоночного столба, 2) грудных и брюшных стенок, 3) нижней челюсти и подъязычной кости относительно дорзального отдела черепа и кожных складок головы 4) поступательные и хватательные конечности. Смотря по эффекту, получающемуся в результате мускульного сокращения, на конечностях принято различать следующие мускулы: а) сгибатели и разгибатели, б) приводящие и отводящие, в) вращатели наружу ивращатели внутрь и г) напрягатели. Эти определения вполне совпадают с теми видами движения, которые уже рассмотрены по отношению ккаждому типу суставов.
Каждая ось движения имеет лишь две мускульные группы, из которых одна действует на сустав в направлении, диаметрально противоположном другой. Отсюда становятся понятными употребляемые по отношению к мускулам названия антагонистов—antagonist—и синергистов, или сотоварищей,—synergetes s. socii. Так, например, разгибатели какого-либо сустава увеличивают его угол, а сгибатели уменьшают, следовательно, разгибатели и сгибатели будут друг другу антагонистами; мускулы, действующие на сустав только в одном направлении, например, сгибая его, будут друг другу синергистами.
Мускул обладает очень совершенной упругостью, вследствие чего он даже в нерабочие моменты находится в слегка растянутом, т. е. напряженном, состоянии (в состоянии мышечного тонуса) и вполне гарантирован от удлинения после своего растяжения действием антагонистов. Эластичность брюшка предохраняет также от порывистых, дергающих движений рычагов при начале сокращения. Этим создается плавная работа на рычагах и устраняется нецелесообразная трата сил на предварительное натяжение самого мускульного брюшка перед сокращением.
Фасции. Фасции—fascia—представляют собой оболочки или пластины, построенные из плотной соединительной ткани. Они формируют для мускулатуры в целом, а также для некоторых других органов дополнительный к скелету фиброзный сложный пластинчатый остов. В его пластины заключены как отдельные мускулы, так и мускульные группы и, наконец, вся мускулатура и сосудисто-нервные пучки.
Общее распределение этого фиброзного остова таково. Все тело животного под кожным покровом одето, как чехлом, подкожной, или поверхностной фасцией—fascia subcutanea s. superficialis. Эта фасция имеет отношение главным образом к подкожным мускулам туловища.
Под поверхностной фасцией лежит второй сложный чехол— глубокая фасция—fascia profunda, —которая уже непосредственно прилегает к поверхностным скелетным мускульным группам, одевая последние в оболочку.
Мускулатуры туловища и хвоста. Среди хордовых животных мускулатура туловища наиболее примитивно построена у ланцетника. За некоторым изъятием она представляет в общем два сильноразвитых, но не дифференцированных мускула — правый и левый. Каждый из них имеет миомерию, т. е. расчленен на десятки (до 60) сегментов поперечными соединительнотканными перегородками. Этасегментация распространяется и на область жаберного аппарата, располагаясь дорзально от него. Короткие пучки мускульных волокон между поперечными перегородками всюду идут исключительно в продольном направлении.
У стоящих выше по зоологической лестнице рыб, одновременно с резким обособлением головы и плавникообразных конечностей мускулатура тела распадается на мускулы головы, туловища, хвоста и конечностей. В мускулатуре туловища и хвоста с такой же ясной миомерией, как и у ланцетника, на каждой симметричной половине появляется уже по продольной боковой перегородке, отъединяющей дорзальную мускулатуру от вентральной. Дорзальный мускул—m. dorsalis,—иннервируемый дорзальными ветвями спинномозговых нервов, в общем напоминает простые отношения у ланцетника, причем краниальный отдел его заканчивается на черепе и дорзальной части плечевого пояса. Вентральный мускул—m. ventralis,— обслуживаемый вентральными ветвями спинномозговых нервов, у некоторых рыб начинает дифференцироваться. Эта дифференциация касается только области туловища, так как хвостовой отдел сохраняет простоту миомерии и продольное направление мускульных пучков.
Изменения на туловище выражаются в выделении у средней сагиттальной линии живота парного прямого мускула—m. rectus—с ясной миомерией; на боковых сторонах (как у ганоидных, так и у костистых рыб) мускульная масса, сохраняя миомерию, расчленяется на два косых пласта—поверхностный и глубокий. Поверхностный пласт представляет собой косой наружный мускул—m. obliquus externus; его пучки идут спереди и сверху назад и вниз. Глубокий пласт в виде косого внутреннего мускула—m. obliquus interims,— пучки которого направлены сзади и сверху вперед и вниз, перекрещивает первый. Сегментальное расчленение остается все же ясно выраженным и по всей вентральной мускулатуре.
У головы рыб из вентральной мускулатуры выделяется висцеральная мускулатура. Она направляется от коракоидной кости к нижней челюсти, подъязычной дуге и жаберным дугам. К челюсти у средней сагиттальной линии идет m. coracomandibularis, рядом с ним лежит идущий к подъязычной кости
Мускулатура рыб. Мускулатура — это специализированная сократительная ткань, которая подразделяется на два основных типа: поперечнополосатую произвольную и гладкую непроизвольную мускулатуру.
Мясо свежей пресноводной рыбы большинства пород содержит влаги—70—80%, белков—17—19%, солей— 1—1,3%, жира—от 0,5 до 10%, у некоторых рыб несколько больше. Содержание жира зависит от возраста рыбы, ее питания и время вылова, рыба осенью имеет больший процент жира, чем весной. У рыб лососевых пород жирность доходит до 20—24%.
 Гладкая непроизвольная мускулатура у рыб работает независимо от волевых актов, хотя тоже иннервирована и подчиняется действию нервных и гормональных стимулов. Гладкая мускулатура способна к спонтанным ритмическим сокращениям даже вне организма.
Гладкая непроизвольная мускулатура у рыб работает независимо от волевых актов, хотя тоже иннервирована и подчиняется действию нервных и гормональных стимулов. Гладкая мускулатура способна к спонтанным ритмическим сокращениям даже вне организма.
Рис.71.Продольное сечение быстрой скелетной мышцы
Например, полный цикл периодического движения желудка и кишечника рыб при комнатной температурре занимает несколько минут. Раздражение ветви, блуждающего нерва, иннервирующей пищеварительный
тракт, вызывает задержку сократительной деятельности. Характеристика деятельности плавательного, мочевого, желчного пузырей рыбы не имеет принципиальных отличий от работы гладкой мускулатуры кишечника. Для работы этих органов не требуется мгновенной реакции и мощного действия. Сокращение гладкой мускулатуры стимулируется многократными нервными и электрическими импульсами. Быстрое растяжение гладких мыщечных волокон под действием внешних сил, например растяжение желудка проглоченной пищей, вызывает ответное сокращение. В стенках полых органов (желудка, кишечника, пузырей) имеются продольные и кольцевые гладкие мышечные волокна. Гладкая мускулатура имеется также в селезенке, радужке глаза и в аппарате движения хрусталика глаза, в стенках кровеносных сосудов и жабрах.
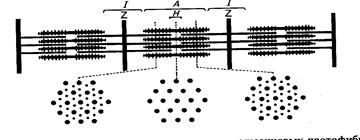 Рис.72. Схема строения попе-
Рис.72. Схема строения попе-
речнополосатой мышцы с перекрывающими друг друга рядами тонких и толстых нитей, с поперечными мостиками и Z пластинками в середине 1-дисков.Вверху- продольный разрез, внизу-поперечный, через разные участки саркомера.
СТРОЕНИЕ И РАБОТА ПОПЕРЕЧНОПОЛОСАТОЙ МУСКУЛАТУРЫ Поперечно-полосатая произвольная мускулатура, или туловищно-хвостовая, является основным двигателем при плавании рыб. Деятельность ее в значительной мере определяется волевыми актами, хотя при плавании большую роль играет автоматизм.
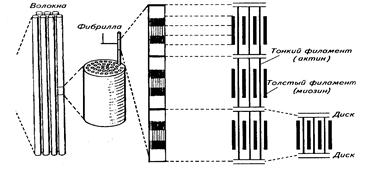
Туловищно-хвостовая мускулатура рыб состоит из отдельных сегментов (миомеров), число которых равно числу позвонков. У рыб вытянутой формы число миомеров около 100 и больше (угорь), у рыб обыкновенной формы (карп, окунь, сельдь, лосось, камбала) примерно 50, у коротких рыб (бычки спинороги, ставрида) около 25 миомеров. Сегментарное строение характерно как для красной, или темной, латеральной мускулатуры, так и для белой, или светлой, составляющей большую часть всей мышечной массы. Несмотря на сегментарное строение, красная мускулатура выступает как единое целое. Все ее мышечные волокна направлены вдоль оси тела. Сокращение этих волокон вызывает латеральные изгибы тела. Белая мускулатура может быть разделена на спинную — выше средней линии тела - и боковую - ниже средней линии тела.
Соединительнотканные элементы, отделяющие спинную мускулатуру от боковой в один миомер, называются миосептами. К ним присоединяются мышечные волокна, с помощью которых сокращение мускулатуры передается костному скелету и вызывает локомоторные волнообразные движения рыбы.
Миомеры имеют сложную геометрическую форму, поэтому поперечный разрез тела рыбы рассекает несколько миомеров, представляющих собой как бы серию вложенных один в другой конусов, толщина которых приблизительно равна длине позвонка. У тунцов и акул таких слоев бывает 17—18. Непосредственно после импульса наблюдается короткий латентный (скрытный) период длительностью в несколько милли секунд. Затем происходят быстрое сокращение и более медленное расслабление. Время, требующееся для полного одиночного цикла, колеблется от 30 до 100 милли секунд.
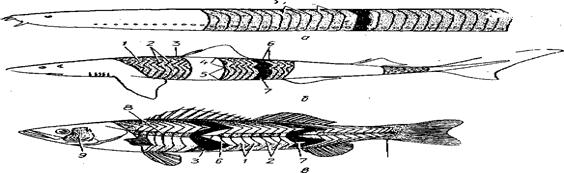
Рис.74. а – миксина; б – акула; в - судак; 1,2- миомеры; 3-миосепты; 4-спинная мускулатура; 5 - латеральная мускулатура; 6-мышечные конусы направленные вперед; 7 - мышечные конусы, направленные назад; 7- поверхностная темная мускулатура; 9-оперкульарно -челюстная группа мышц; 10-группа мышц, сводящих и разводящих лучи хвостового плавника.
Если подавать на мышечное волокно стимулирующие импульсы до конца полного расслабления, то наблюдается продолжительное - изометрическое сокращение, или тетанус. Тетанус бывает частичный и полный. Изометрические напряжения мускулатуры несвойственны рыбам в обычных условиях существования. Они наблюдаются при экспериментальных работах и при электролове. Полный тетанус светлых волокон у ската наступает при стимуляции с частотой 5— 10 Гц, а максимальное напряжение развивается в изометрическом сокращении при частоте 20 Гц. Тетанус быстрых волокон у трески наблюдается при стимуляции с частотой 35—45 Гц, а максимальное напряжение — при частоте стимуляции 250— 400 Гц. По-видимому, гладкий тетанус мышечных волокон наступает при частоте стимуляции, приблизительно в 2—3 раза превышающей максимальную частоту произвольных плавательных движений.
При электрическом раздражении туловищной мускулатуры целой рыбы можно наблюдать тетанус, т. е. судорожное напряжение мускулатуры, но он маскируется множеством нерегулярных подрагиваний. Это явление можно наблюдать, если приложить стимулирующие электроды к боковой поверхности карпа и подавать электрические импульсы с частотой 50 Гц и амплитудой 5 В.
Естественные конвульсивные движения наблюдаются во время нереста у многих видов рыб. Изгибаются - и дрожат во время «брачных игр» лососевые, карповые и некоторые другие рыбы. Титаническое сокращение тела наблюдается при икрометании и выделении молок.
Коэффициент полезного действия плавательной мускулатуры рыб, по всей видимости, не превышает 25%. Коэффициент полезного действия двигательной системы сперматозоидов, как показывают энергетические расчеты, имеет близкую величину — порядка 20%. Это показывает, что сократительные элементы в организме имеют одинаковую физическую и химическую природу.
Мышечная система и электрические органы. Мышечную систему рыб, как и других позвоночных, разделяют на мышечную систему тела (соматическую) и внутренних органов (висцеральную). В первой выделяют мускулы туловища, головы и плавников. Внутренние органы имеют свою мускулатуру. Мышечная система взаимосвязана со скелетом (опора при сокращении) и нервной системой (к каждому мышечному волокну подходит нервное волокно, и каждая мышца иннервируется определенным нервом). Нервы, кровеносные и лимфатические сосуды располагаются в соединительнотканной прослойке мышц, которая в отличие от мышц млекопитающих невелика.
У рыб, как и других позвоночных, сильнее всего развита туловищная мускулатура. Она обеспечивает плавание рыбы. У настоящих рыб она представлена двумя большими тяжами, расположенными вдоль тела от головы до хвоста (большая боковая мышца - m. lateralis magnus) (рис.). Продольной соединительнотканной прослойкой эта мышца делится на спинную (верхнюю) и брюшную (нижнюю) части.
Боковые мышцы разделены миосептами на миомеры, число которых соответствует количеству позвонков. Наиболее отчетливо миомеры видны у личинок рыб, пока их тела прозрачны. Мышцы правой и левой сторон, поочередно сокращаясь, сгибают хвостовой отдел тела и изменяют положение хвостового плавника, благодаря чему тело двигается вперед.
Над большой боковой мышцей вдоль тела между плечевым поясом и хвостом у осетровых и костистых лежит прямая боковая поверхностная мышца (m. rectus lateralis, m. lateralis superficialis). У лососевых в ней откладывается много жира. По нижней стороне тела тянется прямая брюшная мышца (m. rectus abdominalis); у некоторых рыб, например, угрей, ее нет. Между ней и прямой боковой поверхностной мышцей располагаются косые мышцы (m. obliguus).
Группы мышц головы управляют движениями челюстного и жаберного аппаратов (висцеральная мускулатура), плавники имеют свою мускулатуру. Наибольшее скопление мускулов определяет и расположение центра тяжести тела: у большинства рыб он находится в спинной части.
Деятельность туловищных мышц регулируется спинным мозгом и мозжечком, а висцеральная мускулатура иннервируется периферической нервной системой, возбуждаемой непроизвольно.
Различают поперечнополосатые (действующие в значительной степени произвольно) и гладкие мышцы (которые действуют независимо от воли животного). К поперечно - полосатым относятся скелетные мышцы тела (туловищные) и мышцы сердца. Туловищные мышцы могут быстро и сильно сокращаться, однако скоро утомляются. Особенностью строения сердечных мышц является не параллельное расположение обособленных волокон, а разветвление их кончиков и переход из одного пучка в другой, что обусловливает непрерывную работу этого органа.
Гладкие мышцы также состоят из волокон, но гораздо более коротких и не обнаруживающих поперечной исчерченности. Это мышцы внутренних органов и стенок кровеносных сосудов, имеющие периферическую (симпатическую) иннервацию. Поперечнополосатые волокна, а следовательно, и мышцы делят на красные и белые, различающиеся, как следует из названия, цветом. Цвет обусловлен наличием миоглобина - белка, легко связывающего кислород. Миоглобин обеспечивает дыхательное фосфорилирование, сопровождающееся выделением большого количества энергии. Красные и белые волокна различны по целому ряду морфофизиологических характеристик: цвету, форме, механическим и биохимическим свойствам (интенсивность дыхания, содержание гликогена и т. д.). Волокна красной мышцы (m. lateralis superficialis) - узкие, тонкие, интенсивно кровоснабжаемые, расположенные поверхностно (у большинства видов под кожей, вдоль тела от головы до хвоста), содержат в саркоплазме больше миоглобина; в них обнаружены скопления жира и гликогена.
Возбудимость их меньше, отдельные сокращения длятся дольше, но протекают медленней; окислительный, фосфорный и углеводный обмен интенсивнее, чем в белых. В мышце сердца (красной) мало гликогена и много ферментов аэробного обмена (окислительный обмен). Она характеризуется умеренной скоростью сокращений и утомляется медленнее, чем белые мышцы. В широких, более толстых, светлых белых волокнах m. lateralis magnus миоглобина мало, меньше в них гликогена и дыхательных ферментов.
Углеводный обмен происходит преимущественно анаэробно, и количество выделяемой энергии меньше. Отдельные сокращения быстры. Мышцы быстрее сокращаются и утомляются, чем красные. Лежат они более глубоко. Красные мышцы постоянно деятельны. Они обеспечивают длительную и непрерывную работу органов, поддерживают постоянное движение грудных плавников, обеспечивают изгибы тела при плавании и поворотах, непрерывную работу сердца. При быстром движении, бросках активны белые мышцы, а при медленном - красные. Поэтому наличие красных или белых волокон (мышц) зависит от подвижности рыб: "спринтеры" обладают почти исключительно белыми мышцами. У рыб, которым свойственны продолжительные миграции, кроме красных Боковых мышц имеются добавочные красные волокна в белых мышцах.
Основную массу мышечной ткани у рыб составляют белые мышцы. Например, у жереха, плотвы, чехони на их долю приходится 96,3; 95,2 и 94,9% соответственно. Белые и красные мышцы различаются по химическому составу. В красных мышцах содержится больше жира, тогда как в белых мышцах больше влаги и белка. Толщина (диаметр) мышечного волокна изменяется в зависимости от вида рыб, их возраста, величины, образа жизни, а у прудовых рыб - от условий содержания. Например, у карпа, выращенного на естественной пище, диаметр мышечного волокна составляет (мкм): у мальков - 5-19, сеголетков - 14-41, двухлетков - 25-50.
Туловищная мускулатура образует основную долю мяса рыбы. Выход мяса в процентах общей массы тела (мясистость) неодинаков у разных видов, а у особей одного вида различается в зависимости от пола, условий содержания и др. Мясо рыб усваивается быстрее, чем мясо теплокровных животных. Оно чаще бесцветно (судак) имеет оттенки (оранжевый - у лососевых, желтоватый у осетровых и др.) в зависимости от наличия различных жиров и каротиноидов. Основную массу белков мышц рыб составляют альбумины и глобулины (85%), всего же у разных рыб выделяют 4 - 7 фракций белков.
Химический состав мяса (вода, жиры, белки, минеральные вещества) различен не только у разных видов, но и в разных частях тела. У рыб одного вида количество и химический состав мяса зависят от условий питания и физиологического состояния рыбы. В нерестовый период, особенно, у проходных рыб, расходуются резервные вещества, наблюдается истощение и, как следствие, уменьшается количество жира и ухудшается качество мяса.
У кеты, например, во время подхода к нерестилищам относительная масса костей увеличивается в 1,5 раза, кожи - в 2,5 раза. Мышцы оводняются - содержание сухого вещества снижается более чем в два раза; из мышц практически исчезают жир и азотистые вещества - рыба теряет до 98,4% жира и 57% белка. Особенности окружающей среды (в первую очередь пищи и воды) могут сильно изменять пищевую ценность рыбы: в заболоченных, тинистых или загрязненных нефтепродуктами водоемах рыбы имеют мясо с неприятным запахом. Качество мяса зависит и от диаметра мышечного волокна, а также количества жира в мышцах. В значительной мере оно определяется соотношением массы мышечной и соединительной тканей, по которому можно судить о содержании в мышцах полноценных мышечных белков (по сравнению с неполноценными белками соединительнотканной прослойки). Это соотношение изменяется в зависимости от физиологического состояния рыбы и факторов внешней среды.
В мышечных белках костистых рыб на белки приходится: саркоплазмы 20 - 30%, миофибрилл - 60 - 70, стромы около 2%. Все многообразие движений тела обеспечивает работа мышечной системы. Она, главным образом, обеспечивает и выделение тепла и электричества в организме рыбы. Электрический ток образуется при проведении нервного импульса по нерву, при сокращении миофибрилл, раздражении светочувствительных клеток, механохеморецепторов и др.
Электрические органы. Своеобразно измененными мышцами являются электрические органы. Эти органы развиваются из зачатков поперечно-полосатой мускулатуры и расположены по бокам тела рыб. Они состоят из множества мышечных пластинок (у электрического угря их около 6000), преобразованных в электрические пластинки (электроциты), переслаиваемые студенистой соединительной тканью. Нижняя часть пластинки заряжена отрицательно, верхняя - положительно. Разряды происходят под действием импульсов продолговатого мозга. Вследствие разрядов вода разлагается на водород и кислород, поэтому, например, в заморных водоемах тропиков около электрических рыб скапливаются мелкие обитатели - моллюски, рачки, привлеченные более благоприятными условиями дыхания.
Электрические органы могут располагаться в разных частях тела: например, у ската, морской лисицы - на хвосте, у электрического сома - на боках. Генерируя электрический ток и воспринимая силовые линии, искаженные встречающимися на пути предметами, рыбы ориентируются в потоке, обнаруживают препятствия или добычу с расстояния нескольких метров даже в мутной воде. В соответствии со способностью к генерации электрических полей рыб разделяют на три группы:
1. Сильно электрические виды - имеют большие электрические органы, генерирующие разряды от 20 до 600 и даже 1000 В. Основное назначение разрядов - нападение и оборона (электрический угорь, электрический скат, электрический сом).
2. Слабоэлектрические виды - имеют небольшие электрические органы, генерирующие разряды напряжением менее 17 В. Основное назначение разрядов - локация, сигнализация, ориентация (обитающие в мутных реках Африки многие мормириды, гимнотиды, некоторые скаты).
3. Неэлектрические виды - не имеют специализированных органов, но обладают электрической активностью. Генерируемые ими разряды распространяются на 10 – 15 м. в морской воде и до 2м. в пресной воде.
Основное назначение генерируемого электричества - локация, ориентация, сигнализация (многие морские и пресноводные рыбы: например, ставрида, атерина, окунь и др.).
Электрические явления имеют собственное значение или сопровождают многие жизненные процессы, если происходит перемещение ионов, а также имеется несимметричное распределение ионов по разные стороны биологических мембран. Это обусловливает наличие электрических потенциалов и возникновение электрических токов. Электрический ток имеет определенное значение в жизни рыб. Он служит для передачи сигналов в некоторых синапсах, приводит к действию отдельные клеточные механизмы. Электрические токи, генерируемые рыбами, позволяют им общаться друг с другом, а также используются для поиска, локации, защиты и нападения.
Природа биотоков и биопотенциалов. Клеточная мембрана отделяет внутреннюю среду клетки, богатую ионами калия, от межклеточной жидкости, богатой ионами натрия. Она окружена снаружи электростатически фиксированными ионами натрия, изнутри - слоем ионов хлора, которые притянуты электростатическими силами к ионам натрия. Ионы калия оттеснены в глубь клетки. Наличие ионной оболочки из положительно заряженных ионов натрия снаружи клетки и ионной подкладки из отрицательно заряженных ионов хлора изнутри мембраны, создает электрическую поляризацию мембраны с разностью потенциала 60—70 мВ в нервных клетках, около 80 мВ в мышечных клетках и 20 мВ в эпителиальных клетках.
Эта поляризация и электрический потенциал поддерживаются низкой проницаемостью мембран для ионов и постоянной работой ионных насосов. Клеточная мембрана обладает высоким электрическим сопротивлением — порядка 1000 Ом/см2 и электрической емкостью около 1 мкФ/см2. Движение ионов по градиенту концентрации и работа ионных насосов порождают ток покоя (рис.). Токи действия возникают при проведении нервного импульса по нерву при сокращении миофибриллы, при работе клетки почечного канальца, энтероцита и т. д. Специфические электрические явления возникают при освещении светочувствительных клеток, раздражении механорецепторов.
При возбуждающем воздействии на клетку потенциал ее мембраны изменяется. Например, при раздражении нервной или мышечной клетки ее мембрана деполяризуется, т. е. мембранный потенциал уменьшается, а затем возвращается к норме через фазу гиперполяризации. Весь процесс занимает тысячные доли секунды. Сложение токов действия отдельных клеток организма рыб, вызывает возникновение так называемых биотоков – токов возникающих в живом организме, которые обнаруживаются даже на некотором расстоянии от места их возникновения. Запись биотоков работающей мускулатуры называют электромиограммой. По электромиограммесудят о том, как, когда и какие группы мышц работают при различных двигательных актах. Биотоки регистрируются при плавании рыб дыхательных движениях. Электрокардиограмма рыб имеет характерные «зубцы» PQRST. Перед зубцом Р, являющимся результатом сокращения предсердия, у рыб имеется волна возбуждения венозного синуса. Зубец R сопровождает сокращение желудочка, а зубец Т возникает при электрической переполяризации поверхности желудочка.
Для обнаружения тока действия (акционнъй ток), один электрод надо приложить к невозбужденному участку ткани, а другой — к возбужденному и соединить их с регистрирующим прибором.
Записи биотоков сетчатки глаза (ретины) называются ретинограммами. Характерной особенностью их является то, что волна тока действия в них в отличие от мышц и нервов является волной гиперполяризации, а не волной деполяризации. При освещении колбочек и палочек разность потенциалов между внутренней и внешней поверхностями мембраны растет. По характеру сигналов от сетчатки можно определить спектральную чувствительность.
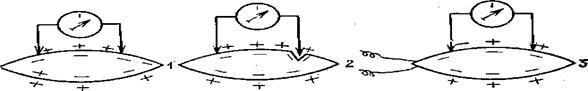
Рис. 74. Мышечные токи:
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 1588; Нарушение авторских прав?; Мы поможем в написании вашей работы!