КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физиология центральной нервной системы 1 страница
|
|
|
|
Ток покоя; 2.— ток повреждения; 3 — ток действия
Запись биотоков головного мозга называется энцефалограммой. Как и у других позвоночных животных, энцефалограммы рыб имеют характер коротких скачков и медленных волн. Их изучение может служить для определения действия медикаментов и токсикантов, а также для решения многих научных задач, связанных с изучением нервной деятельности рыб.
При повреждении ткани в участке повреждения анионы выходят на поверхность, и этот участок становится заряженным отрицательно по отношению к другим участкам. Таким образом, если поставить один электрод на поврежденный (отрицательный.) и второй на неповрежденный (положительный) участки, то регистрирующий прибор покажет наличие тока — тока повреждения.
Биотоки возникают при работе органов и тканей у разных животных, но только у рыб имеются специализированные органы генерирующих электрический ток.
Строение электрического органа. Электрические органы представляют собой столбики, состоящие из клеток плоской формы, — электрические пластинки, расположенные по бокам тела. Толщина этих клеток не превышает 10 мкм. Электрические клетки у большинства рыб имеют мышечное происхождение, у электрического сома они происходят из клеток кожи, а у рыб из семейства стернархид — из нервных клеток. Каждую клетку покрывает оболочка — электролемма.
Форма клеток разнообразна: округлая и цилиндрическая (у гимнотид), розеткообразная (у электрического сома) чашеобразная и дискообразная (у скатов), а сторона пластинки к которой подходят нервные окончания, называется лицевой и под обыкновенным микроскопом выглядит гладкой, а под электронным видны канальцы, по которым проходят нервные волокна.

Рис. 75. Расположение электрических органов у рыб: а – скат; б – японский скат; в – электрический сом; г - электрический угорь; д- нильский слоник; е – нильская щука.
Благодаря наличию полупроницаемой клеточной мембраны происходит перераспределение ионов натрия, калия, кальция, хлора. С противоположной стороны клетки подходят кровеносные сосуды, здесь же располагаются и ядра. Эта сторона поверхности у большинства рыб губчатая, морщинистая, и ее называют нутритивным (питательным) слоем.
У электрических рыб различают два вида мембран: возбудимые при помощи химических медиаторов и возбудимые не только химическим, но и электрическим путем. Мембраны первого вида имеются у таких морских рыб, как электрические скаты, звездочеты и др. мембраны второго вида встречаются у пресноводных рыб.
Неровная поверхность пластинки при возбуждении является электроотрицательно по отношению к ее обратной стороне. Исключение составляет электрический сом. Если поставить электроды на разные точки одной стороны пластинки, то регистрирующий прибор не покажет наличия тока, т. е. лицевая сторона работает как единое целое. Если один электрод поставить на лицевую сторону пластинки, а другой погрузить в ее цитоплазму или поставить на противоположную сторону, то возникает ток напряжением 90—150 мВ.
Контрольные вопросы.
1.Мофологические особенности плавников костистых и хрящевых рыб?
2.Функциональное значение плавников для жизнедеятельности рыб?
3.Каков механизм движения рыб при плавании?
4.Охарактеризуйте механизм полета рыб.
ГЛАВА 11
Центральная нервная система выполняет в организме функции первостепенной важности. Во-первых, она обеспечивает точную регуляцию всех процессов внутри организма, их координацию и интеграцию, благодаря чему все органы и системы действуют согласованно, а организм представляет собой единое функциональное целое. Во-вторых, ей принадлежит функция связи организма с постоянно меняющейся внешней средой.
11.1 Общая физиология центральной нервной системы. Центральная нервная система позвоночных построена из громного количества нейронов, вступающих в тесную связь друг с другом. Однако каждый нейрон относительно самостоятелен. Это обусловливается тем, что протоплазма одной клетки и ее отростков не проникает внутрь другой клетки. Аксон одной нервной клетки, как правило, только соприкасается с дендритами или телом другой. Место контакта двух нейронов называют синапсом. В центральной нервной системе имеется бесчисленное множество синапсов.
В функциональном отношении нейроны делят на три основных класса: 1) чувствительные, или афферентные, 2) двигательные, или эфферентные, 3) вставочные, или промежуточные.
Афферентные нейроны проводят возбуждение от рецепторов в центральную нервную систему, их называют также рецепторными. Тела этих нейронов расположены вне центральной нервной системы и находятся в спинномозговых или черепномозговых ганглиях. Данные нейроны отличаются от других наличием двух длинных отростков — собственно аксона, передающего возбуждение от тела клетки в центры спинного мозга или мозгового ствола, и аксоноподобного дендрита, уходящего на периферию в виде афферентного волокна и ветвящегося там на чувствительные нервные окончания — рецепторы. К рецепторным нейронам относятся также некоторые нейроны в центральной нервной системе, которые получают возбуждение не непосредственно от рецепторов, а через другие истинно рецепторные нейроны. К ним относятся, например, нейроны зрительных бугров.
Рецепторные нейроны называют также чувствительными, или сенсорными, так как они доставляют в центральную нервную систему импульсы, вызывающие различные ощущения.
Эфферентные, или эффекторные, нейроны передают возбуждение из центральной нервной системы к рабочим органам - эффекторам. От тел этих нейронов возбуждение идет на периферию по длинным аксонам. Те эфферентные нейроны, которые посылают импульсы к скелетным мышцам, называются двигательными нейронами, или мотонейронами. Их тела лежат в вентральных рогах спинного мозга, в продолговатом и среднем мозге.
Многие эфферентные нейроны передают импульсы не прямо на периферию, а через другие, ниже расположенные нейроны. Например, эфферентные нейроны среднего мозга посылают импульсы к мотонейронам спинного мозга. Эфферентные нейроны вегетативной нервной системы находятся вне центральной нервной системы - в периферических ганглиях.
Вставочные (промежуточные, контактные нейроны, интернейроны) образуют самую многочисленную группу нейронов в центральной нервной системе. Они осуществляют связь между рецепторными и эффекторными нейронами. По характеру вызываемого ими эффекта контактные нейроны делят на возбуждающие и тормозящие.
11.2Строение и классификация межнейронных синапсов. Межнейронным, или центральным, синапсом называют место контакта окончания аксона с телом или отростками другой нервной клетки. Если аксон оканчивается на теле (соме) другой нервной клетки, синапс будет аксосоматическим, на дендритах - аксодендритическим, на аксонах- аксоаксональным. Синапсы состоят: 1) из пресинаптической мембраны, покрывающей окончание аксона (синаптическую бляшку), 2) синаптической щели шириной примерно 200—ЗООА и 3) постсинаптической мембраны соседней нервной клетки. Внутри синаптической бляшки находится много пузырьков, в которых, как полагают, содержится вещество, обеспечивающее передачу импульса через синапс. Это вещество получило название медиатора. Кроме пузырьков медиатора, имеются митохондрии — специфические структуры, поставляющие клетке энергию. По функциональному признаку различают возбуждающие и тормозящие, или тормозные, синапсы. Возбуждающие и тормозящие синапсы имеют некоторые структурные отличия. Так, возбуждающие синапсы большей частью являются аксодендритическими, характеризуются относительно широкой синаптической щелью (примерно ЗООА), толстой, плотной постсинаптической мембраной. В синаптической щели могут быть специальные включения в виде пластинок внеклеточного вещества. Пузырьки медиатора относительно крупные, округлой формы. Тормозные синапсы имеют более узкую синаптическую щель (в 1,5—2 раза уже, чем возбуждающие); постсинаптическая мембрана более тонкая; в синаптической щели нет включений внеклеточного вещества; синаптические пузырьки овальной формы, по размеру меньше, ряд пузырьков уплощен.
Природа медиаторов возбуждающего и тормозного синапсов также различна. Химическим передатчиком возбуждения в возбуждающих синапсах является ацетилхолин; полагают, что такими свойствами обладает и глутаминовая кислота. В возбуждающих центральных синапсах медиаторами служат адреналин, норадреналин, 5-окситриптамин (близкие по строению вещества), а также гистамин.
К медиаторам, выделяющимся в тормозящих синапсах, относят аминокислоту глицин. Г лицина в участках спинного мозга, где распола
-гаются тормозные нейроны Реншоу, значительно больше, чем в других соседних местах. Кроме того, когда методом электрофореза к телу нерв
ной клетки, подводили глицин, возникала - гиперполяризация клеточной мембраны, что является доказательством постсинаптического торможения. Вторым тормозящим медиатором считают гамма-аминомаслянуго кислоту (ГАМК). Этот медиатор был открыт у ракообразных. Затем он был найден в головном мозге в частности в окончаниях тормозящих нейронов. Механизм передачи возбуждения в синапсах центральной нервной системы.Создание тонких методов регистрации импульсов отдельных нейронов с помощью высокочувствительных электрических приборов произвело переворот в исследованиях передачи импульсов через синапс. Этот метод заключается в том, что в нервную клетку осторожно вводят очень тонкую стеклянную пипетку, заполненную солевым раствором, проводящим электрический ток (например, концентрированным раствором хлористого калия). Если ввести осторожно пипетку и закрепить ее неподвижно, клеточная оболочка быстро стягивается вокруг кончика пипетки, и такие нервные клетки способны нормально функционировать в течение нескольких часов.
Механизм передачи возбуждения заключается в следующем. Под действием приходящих к окончанию аксона нервных импульсов происходит выделение из синаптических пузырьков молекул медиатора ацетилхолина. Они диффундируют через пресинаптическую мембрану в синаптическую щель, а затем к постсинаптической мембране и прикрепляются к определенным ее участкам, называемым рецепторными. Здесь медиатор взаимодействует с ее структурными компонентами—белково-липоидными комплексами. В результате происходит кратковременное увеличение диаметра пор в мембране (в 3—4 раза), что приводит к сильному повышению проницаемости постсинаптическоймембраны для ионов натрия, которые устремляются из синаптической щели через постсинаптическую мембрану. Происходит деполяризация постсинаптической мембраны и возникает возбуждающий постсинаптический потенциал (ВПСП), и \когда он достигает критического уровня, появляется потенциал действия. Выделившийся ацетилхолин очень быстро разрушается ферментом холинэстеразой на холин и уксусную кислоту. Поэтому каждая выделившаяся порция медиатора действует кратковременно, успевая вызвать только один импульс. Сразу после, возникновения импульса происходит уменьшение диаметра пор в постсинаптической мембране, и ионы натрия, которые в гидратированном состоянии крупнее ионов калия, перестают переходить внутрь клетки, а ионы калия свободно выходят наружу в синаптическую щель. Благодаря этому всего лишь за милли секунду или даже быстрее восстанавливается исходная полярность постсинаптической мембраны и включается натриево-калиевый насос (рис. 76).
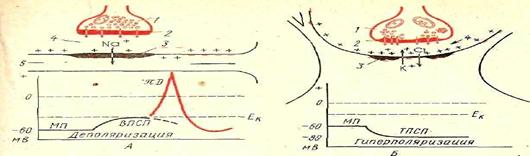
Рис76. Схема функции синапса:
А - возбуждающего; Б - тормозящего; 1 - пузырьки медиатора; 2 - лресинаптическая мембрана; 3 — постсинантическая мембрана; 4 - синаптическая щель; Ек - критический уровень деполяризации; МП - местный потенциал; ВПСП - возбуждающий и ТПСП — тормозной посчеинаптические потенциалы; ПД — шненциал действия.
11.3Учение о рефлексе. Основной формой деятельности центральной нервной системы является рефлекс.
Рефлексом называют реакцию организма на раздражение рецепторов, осуществляемую при участии центральной нервной системы.
Учение о рефлексе имеет свою историю, охватывающую столетия. Основоположником представлени о простейших рефлекторных актах является французский философ и естествоиспытатель Рене Декарт(1595—1650). Он описал такие автоматические действия, как отдергивание руки при прикосновении к огню, подмигивание глаз при раздражении роговицы и т. д. Большую роль в дальнейшем развитии учения о рефлексе сыграл чешский анатом и физиолог Георг Прохаска, который и ввел в науку этот термин. Слово «рефлекс» в переводе с латинского значит «отражение», т. е. в него вкладывался такой смысл, что всякое действие организма является отражением какого-либо воздействия на него и для каждого действия имеется своя причина. Представление о рефлексе как о явлении, впервые сделал русский ученый-материалист И. М. Сеченов в своей знаменитой книге «Рефлексы головного мозга» (1863), обосновавший положение о том, что «все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы». И. П. Павлов, развивая и продолжая идеи И. М. Сеченова, создал учение об условных рефлексах и применил метод условных рефлексов для исследования функций головного мозга.
Понятие о рефлексе и рефлекторной дуге. Морфологической единицей нервной системы считается нейрон, а функциональной единицей нервной деятельности является рефлекс. Рефлекторные акты представляют собой основную специфическую функцию нервной системы.
Прежде чем рассматривать деятельность отдельных частей центральной нервной системы, необходимо рассмотреть основные закономерности рефлекторных реакций.
Под рефлексом понимают непроизвольную ответную реакцию организма на раздражение, осуществляемую центральной нервной системой и выражающуюся мышечной или секреторной деятельностью.
Любая рефлекторная реакция осуществляется через так называемую рефлекторную дугу, которая состоит из двух и более нейронов (рис.77а). Рефлекторная дуга это путь, по которому проходят нервные импульсы от рецептора до эффектора. Сущность рефлекторного акта состоит в последовательном возбуждении различных участков рефлекторной дуги. Раздражение внешней среды воспринимает рецептор, т.е.окончания центростремительного нерва в коже (экстерорецептор), в слизистых оболочках (интерорецептор) и в других частях организма.
Возникший в результате раздражения рецептора процесс возбуждения передается через центростремительный (афферентный) нерв в чувствительную клетку, а от неё, через дендрит,- двигательной клетке серого вещества спинного мозга. Далее импульс возбуждения идёт по центробежному (эфферентному) нерву к рабочему органу - эффектору,т. е. к мышце или железе.
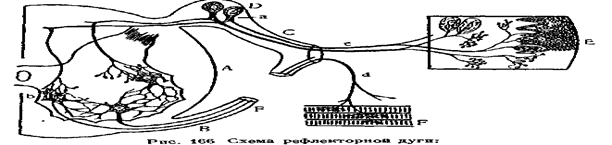
Рис.76 а
А — спинной мозг; В — двигательный (нижний) корешок, С—чувствительный (верхний) корешок; D — спинномозговой узел; Е—кожа, F—мышца скелета, о — клетки чувствительных нейронов в спинномозговых узлах, Ь — клетки двигательных нейронов в передних рогах, с — периферические отростки чувствительных нейронов, которые, оканчиваясь в коже, образуют рецепторы d—нейрит двигательных нейронов, оканчивающихся в скелетных мышцах.
В осуществлении рефлекторной функции может участвовать вся центральная нервная система.
Необходимым условием рефлекторного процесса является целость всех составляющих частей дуги. Нарушение одной из этих частей, например, рецептора, делает невозможным осуществление рефлекса через данную дугу.
Для животных и рыб рефлекторные акты имеют большое приспособительное значение. Выключение некоторых рефлекторных функций центральной нервной системы может значительно нарушить отправление важнейших для жизни частей организма.
Рефлекторные акты осуществляются через нервные клетки различных этажей центральной нервной системы. При этом возбуждение по многочисленным проводящим путям способно широко распространяться в центральной нервной системе. Рефлекс — это сложная реакция организма, осуществляемая нервными центрами. Поэтому необходимо обратить внимание на некоторые основные свойства центров, от которых в значительной степени зависит конечный эффект рефлекторного акта. Основные свойства нервных центров Одностороннее проведение. У позвоночных животных импульс в центральной нервной системе проводится только в одном направлении: от рецепторного нейрона через промежуточный к эффекторному. Такая направленность нервного процесса, характерная для рефлекторной дуги, обусловлена односторонним проведением возбуждения в синапсах. Такое явление можно объяснить тем, что синапс представляет собой односторонне проницаемую перепонку.
Замедленное проведение. Проведение возбуждения в центральной нервной системе совершается значительно медленнее, чем по нервам. Это объясняется свойствами синапсов задерживать проведение возбуждений с одного нейрона на другой. Проведение через синапс у лягушки осуществляется в течение 0,001 —0,003 секунды. Очевидно, чем большее число нейронов входит в данную рефлекторную реакцию, тем больше затрачивается времени на её осуществление.
Временем рефлекса называется период, протекающий от момента раздражения до начала реакции. Например, время рефлекса моргания у человека составляет 0,05—0,2 секунды.
Облегчение. Раздражение, приходящее в нервные центры, не исчезает сразу, а оставляет след в виде повышенной возбудимости блуждающего нерва.
Торможение. В центральной нервной системе, наряду с возбуждением, встречается другой процесс — торможение. Благодаря тесной функциональной взаимосвязи возбудительного и тормозного процесса, а также взаимодействия между отдельными нейронами, называемого координацией, обеспечивается наиболее точное и тонкое мышечное движение и приспособление тех или иных двигательных актов к тем или иным воздействиям внешней среды.
Впервые процесс центрального торможения был открыт Сеченовым. Для возникновения торможения необходимо одновременное поступление в центральную нервную систему двух или больше импульсов возбуждения по различным рефлекторным путям. Более сильное раздражение вызывает угнетение аффекта от более слабого раздражения. Объясняется это тем, что в центральной нервной системе происходит конфликт между возбуждениями, и одно из них подавляет другое. Так, например, раздражение серого вещества головного мозга может повести к задерживанию спинных рефлексов и т. д. Природа процесса торможения ещё не ясна.
Рефлекторное сокращение мышц сгибателей ведёт к расслаблению мышц разгибателей той же конечности. Мышцы, производящие сгибание конечности в одном или нескольких суставах, называются синергистами, а мышцы, производящие разгибание,т.е. противоположное действие, называются антагонистами.
Взаимоотношения между мышцами антагонистами в период любого двигательного акта обусловлены соотношением нервных процессов в центрах. Возбуждение центров антагонистических групп мышц возникает в момент сокращения и обусловлено восприятием раздражения рецепторами, заложенными в мышцах и сухожилиях; эти рецепторы называются проприорецептор а м и.
По имеющимся данным, возбуждение центра одной группы мышц связано с торможением центра другой, антагонистической, мускульной группы (так называемый принцип взаимносопряжённой, реципрокной иннервации). Реципрокные соотношения между центрами осуществляются в широких пределах. Так, например, возбуждение центра сгибания одной конечности вызывает не только торможение центра разгибания этой же конечности, но и торможение центра сгибания, и возбуждение центра разгибания мышц другой конечности. Вследствие этого, при сгибании одной конечности происходит разгибание другой.
Реципрокные взаимоотношения имеются также между центрами передних и задних конечностей. В центрах мускулатуры конечностей, участвующих при двигательных актах, постоянно происходит перераспределение возбуждения и торможения.
г) Изменчивость координации и доминанта
Изучение взаимоотношений между центрами, иначе — координационных отношений, показывает, что имеются относительно постоянные, сложившиеся в процессе фило- и онтогенетического развития явления координации, как, например, реципрокная иннервация антагонистических мышц. Но в нервной системе мы наблюдаем и изменчивость явлений координации. Координационные отношения складываются в результате целого ряда внутренних и внешних условий к моменту возникновения данного раздражения. Ответная рефлекторная реакция может поэтому значительно видоизменяться.
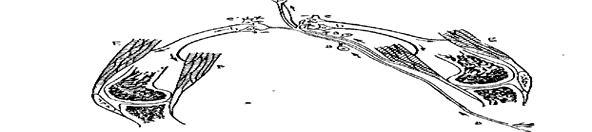
Рис.77. Схематическое изображение реципрокной
иннервации.
Раздражение чувствительного нерва вызывает на соответствующей стороне возбуждение двигательного нейрона, принадлежащего сгибателю- F; одновременно вызывается торможение двигательного нейрона - е, принадлежащего разгибателю, раздражение на другой стороне производит противоположный эффект.
В связи с динамичностью координационных процессов особый интерес представляет установленный Ухтомским принцип доминанты, позволяющий понять механизм преобладания одних очагов возбуждения над другими.
Доминанта — это господствующий очаг возбуждения в центральной нервной системе. Появляется такой очаг при условии повышения возбудимости какого-либо центра рядом волн возбуждения, оптимальных по ритму и силе или по гуморально - химическим влияниям. Доминантный очаг возбуждения привлекает к себе возбуждения, приходящие к другим нервным центрам, усиливается за счёт этих возбуждений и затормаживает все остальные центры.
11.4 Морфология и функциональная характеристика центральной нервной системы. Центральная нервная система рыб, как и у ланцетника, имеет вид трубки. Ее задний отдел - спинной мозг - расположен в канале позвоночника, образованном верхними дугами позвонков. От спинного мозга между каждой парой позвонков вправо и влево отходят нервы, управляющие работой мышц тела и плавников. По нервам от чувствительных клеток на теле рыбы в спинной мозг поступают сигналы о раздражении. Передняя часть нервной трубки рыбы и других позвоночных животных видоизменена в головной мозг, защищенный костями черепной коробки. В головном мозге позвоночных различают отделы: передний мозг, промежуточный мозг, средний мозг, мозжечок и продолговатый мозг. Все отделы головного мозга имеют большое значение в жизнедеятельности рыбы. Например, мозжечок управляет координацией движений и равновесием животного. Продолговатый мозг постепенно переходит в спинной мозг. Он играет большую роль в управлении дыханием, кровообращением, пищеварением и другими важнейшими функциями организма
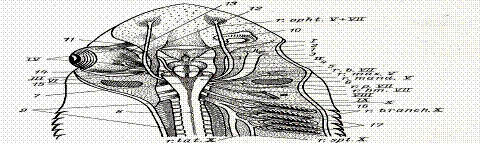
Рис.79. Головной мозг и головные нервы колючей акулы (со спинной стороны, правый глаз удален):
1- предний мозг, 2 – обонятельная доля переднего мозга, 3 – промежуточный мозг, 4 – средний мозг, 5 - мозжечок, 6 – продолговатый мозг, 7 – ромбовидная ямка, 8 – спинной мозг, 9 – спинномозговые нервы, 10 – обонятельный мешок, 11 – верхняя косая мышца глаза, 12 – верхняя прямая мышца глаза, 13 -внутренняя прямая мышца глаза, 14 - наружная прямая мышца глаза, 15 - брызгальце, 16 – первая жаберная щель, 17- вторая – пятая жаберные щели; I-обонятельный тракт; I I – зрительный нерв; III – глазадвигательный нерв; IV- блоковый нерв;V-тройничный нерв;верхнечелюстная ветвь; нижнечелюстная ветвь; VI -,VII-, VIII-,IX-, X-, X -,X -
Нервная система делится на центральную, к которой относятся головной и спинной мозг, и периферическую – нервы, отходящие от головного и спинного мозга. От мозга отходят нервные волокна, окончания которых выходят на поверхность кожи и образуют у большинства рыб ярко выраженную боковую линию, проходящую от головы до начала лучей хвостового плавника. Боковая линия служит для ориентации рыбы:определения силы и направления течения, наличия подводных предметов 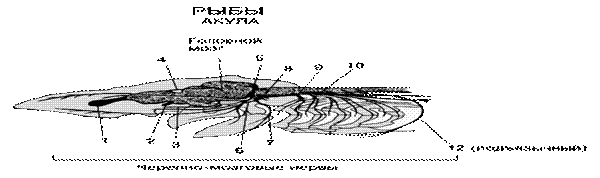
Рис.79. Расположение черепномозговых нервов акулы
Наблюдая поведение окуня в аквариуме, можно заметить, что ответы на раздражение у окуня могут проявляться двояко. Если к окуню прикоснуться, он моментально метнется в сторону. Столь же быстр его ответ на вид пищи. Жадный хищник, он стремительно кидается на свою добычу (мелких рыб и различных беспозвоночных - ракообразных, червей). При виде добычи возбуждение идет по зрительному нерву в центральную нервную систему окуня и сейчас же по двигательным нервам возвращается от нее к мускулам. Окунь плывет к жертве и захватывает ее.
Механизм подобных ответов организма на раздражение врожденный. А такие рефлексы называют безусловными. У всех животных одного вида безусловные рефлексы одинаковы. Они врожденны и передаются по наследству.
Если кормление рыб в аквариуме сопровождается какими-либо действиями (условиями), например зажиганием лампочки или постукиванием по стеклу, то через некоторое время такой сигнал начинает привлекать рыб сам по себе, без подкормки. На подобные сигналы у рыб вырабатываются условные рефлексы, то есть рефлексы, возникающие при определенных условиях. В отличие от инстинктивных безусловных рефлексов условные рефлексы не передаются по наследству. Они индивидуальны и вырабатываются в течение жизни животного.
Головной мозг: У акулы головной мозг состоит из пяти отделов. Передний мозг (telencephalon) крупный, нечетко разделен на правую и левую (полушарии). В передней части полушарии хорошо заметны сильно развитые обонятельные части.
Задняя часть переднего мозга незаметно переходит в промежуточный мозг (diencephalon), который на препарате виден, как углубление между вздутиями переднего и среднего мозга. Средний мозг (mesencephalon) имеет вид крупных парных вздутий (зрительные доли), которые прикрывают собой остальную массу этого отдела мозга.
Четвертый отдел мозга – мозжечок (cerebellum, metencephalon) у акуловых рыб сильно развит и спереди налегает на средний, а сзади на продолговатый мозг.
Последний отдел головного мозга – продолговатый мозг (myelencephalon). Сверху в центральной части продолговатого мозга отчетно видна ромбовидная ямка, представляющая собой полость этого отдела мозга (четвертый желудочек). Она затянута сосудистой оболочкой, которая при препарировании обычно разрушается.
Головные нервы. Характер отхождения головных нервов у акуловых рыб типичен для всех позвоночных животных. От головного мозга у акуловых рыб отходят десять пар головных (черепных) нервов, каждая пара симметрична с обеих сторон головного мозга. Головные нервы обычно имеют двойное обозначение: порядковый номер и название.
I. Обонятельный нерв (nervus olphactorius). У акуловых рыб, как и у всех позвоночных, от переднего мозга отходят выросты его лобных долей – обонятельные тракты. Они образуют у обонятельных капсул расширение – обонятельные луковицы. Обонятельный нерв представляет комплекс коротких чувствительных волокон, идущих от эпителия обонятельного мешка к обонятельной луковице.
II. Зрительный нерв (n. opticus) фактически это зрительный тракт, образующийся как вырост стенки мозга он отходит ниже боковой поверхности промежуточного мозга и пройдя отверстие в стенке глазницы, входит в глазное яблоко, распространяясь по внутренней поверхности сетчатки. Нерв чисто чувствительный.
У всех позвоночных животных движения глазного яблока осуществляется при помощи глазных мышц, каждая из которых одним концом прикрепляется к стенке глазницы, а другим к определенному участку поверхности глазного яблока. От передней части стенки глазницы, отходят 2 косые мышцы глаза; верхняя (m. Obliquus superior) и нижняя (m. Obliquus interior). От задней стенки глазницы пучком отходят 4 прямые мышцы глаза: прямая нижняя (m. Recfus inferior), прямая внутренняя (m. Rectus internus), прямая верхняя (m. Rectus zuperior) и прямая наружная (m. Rectus externus). Эти мышцы иннервируются тремя парами головных нервов – III, IV u VI все эти нервы чисто двигательные.
Глазодвигательный нерв (n. oculomotorius). Отходит от дна среднего мозга, пронизывает стенку черепа и делится у основания прямых мышц глаза на ветви, идущие к четырем мышцам к нижней косой и к нижней внутренней и верхней прямым.
Блоковый нерв (n. trochlcaris). В виде тонкой нити отходит от задне-верхней части среднего мозга, проходит вперед по крыше среднего моста, и проидя через переднюю стенку глазницы, разветвляется в верхней косой мышце глаза. Все остальные головные нервы отходят от продолговатого мозга.
Отводящий нерв (n. abolucens). От дна продолговатого мозга сразу уходит в дно черепной коробки. Он иннервирует наружную прямую мышцу глаза.
Тройничный нерв (n. trigeminus). Это нерв сложный. Он отходит толстым корнем от переднебоковой поверхности продолговатого мозга и сразу же делится на три ветви глазничную, верхнечелюстную и нижнечелюстную. Глазничная ветвь (ramus ophtholmicus) вместе с глазничной ветвью лицевого нерва проходит через глазницу и ветвится в передней части рыла, иннервируя кожные органы чувств.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 1660; Нарушение авторских прав?; Мы поможем в написании вашей работы!