КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Органы слуха у рыб 1 страница
|
|
|
|
Любой источник звука, находящийся на субстрате, помимо излучения классических звуковых волн, распространяющихся в водной или воздушной среде, рассеивает часть энергии, в форме различного рода вибрации, распространяющихся в субстрате и по его поверхности.
Под слуховой системой мы понимаем рецепторную систему, способную воспринимать тот или иной компонент звукоизучения, локализовать и оценивать характер источника, создавал предпосылки для формирования специфических поведенческих реакции организма.
Слуховую функцию у рыб осуществляют, помимо основного органа слуха еще и боковая линия, плавательный пузырь, а также специфические нервные окончания.
Органы слуха рыб развивались в водной среде, которая проводит звук в 4 раза быстрее и на большие расстояния, чем атмосфера. Диапазон восприятия звуков у рыб существенно шире, чем у многих наземных животных и людей.
В жизни рыб слух играет очень большую роль, особенно рыб, обитающих в мутной воде. В боковой линии рыб обнаружены образования, которые регистрируют акустические и другие колебания воды.
Слуховой анализатор человека воспринимает колебания - с частотой от 16 до 20000 Гц. Звуки с частотой ниже Гц относятся к инфразвукам, а выше 20000 Гц - к ультразвукам. Наилучшее восприятие звуковых колебаний наблюдается в интервале от 1000 до 4000 Гц. Спектр звуковых частот, воспринимаемых рыбами, по сравнению с человеком значительно сокращен. Так, например карась воспринимает звуки в диапазоне 4 (31—21760 Гц, карликовый сомик —60—1600 Гц, акула 500- 2500 Гц.
Органы слуха рыб обладают способностью к адаптации факторам окружающей среды в частности, постоянному или однообразному и часто повторяющемуся шуму, например работе землечерпалки, рыба быстро привыкает и шума ее не боится. Также не ухудшает клева рыбы шум проходящего парохода, поезда и даже люди, купающиеся довольно близко к месту ловли, не распугивают рыб. Испуг у рыбы очень кратковременен. Удар блесны о воду, если он произведен без сильного шума, не только не пугает хищника, а возможно настораживает его в ожидании появления чего-либо съедобного для него. Рыба, может воспринимать отдельные звуки, если они вызывают колебания водяной среды. Благодаря плотности воды звуковые волны хорошо предаются через кости черепа и воспринимаются органами слуха рыбы. Рыбы могут слышать шаги человека, идущего по берегу, звон колокольчика, выстрел.
Анатомически, как и у всех позвоночных, основной орган слуха - ухо является парным органом и составляет единое целое с органом равновесия. Отличие заключается только в том, что у рыб нет ушных раковин и барабанных перепонок, так как они живут в другой среде. Орган слуха и лабиринт у рыб одновременно является органом равновесия, он расположен в задней части черепа, внутри хрящевой, или костной, камеры, состоит из верхнего и нижнего мешочков, в которых находятся отолиты- камешки.
Орган слуха рыб представлен только внутренним ухом и состоит из лабиринта. Внутреннее ухо стато - акустический парный орган. У хрящевых рыб состоит из перепончатого лабиринта заключенного в хрящевую слуховую капсулу - боковое расширение хрящевого черепа позади глазницы. Лабиринт, представлен тремя перепончатыми полукружными каналами и тремя отолитовыми органами — утрикулюсом, саккулюсом и лагеной (рис. 91,92,93). Лабиринт делится на две части: верхнюю, к которой относятся полукружные каналы и утрикулюс, и нижнюю часть—саккулюс и лагена. Три изогнутые трубочки полукружные каналы лежат в трех взаимно перпендикулярных плоскостях и концами открываются в преддверье или перепончатый мешок. Он подразделяется на две части верхнюю овальный мешочек и более крупную нижнюю - круглый мешочек, от которого отходит небольшой вырост - лагена.
Полость перепончатого лабиринта заполнена эндолимфой, в которой взвешены мелкие кристаллики отоконии. В полости круглого мешочка обычно находятся более крупные известковые образования отолиты состоящие из соединений кальция. Колебания, которых воспринимаются слуховым нервом. Окончания слухового нерва подходят к отдельным участкам перепончатого лабиринта, покрытым чувствующим эпителием – слуховым пятнам и слуховым гребням. Звуковые волны передаются непосредственно через ткани восприятия колебания, которые воспринимаются слуховым нервом.
Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях. Каждый полукружный, канал впадает в утрикулюс двумя концами, один из которых - расширяется в ампулу. Здесь имеются возвышения, называваемые слуховыми пятнами, или макулами, где расположены скопления чувствительных волосковых клеток. Тончайшие волоски этих клеток соединены студнеобразным веществом, образуя купулу. К волосковым клеткам подходят окончания VIII пары-черепно-мозговых нервов.
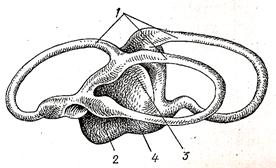
| Рис.91.Схема перепончатого лабиринта акулы: 1-полукружные каналы, 2-овальный мешочек, 3-круглый мешочек, 4-лагена |
В утрикулюсе костистых рыб находится один крупный отолит. Отолиты располагаются также в лагене и саккулюсе. Отолит саккулюса используется для определения возраста рыб. Саккулюс хрящевых рыб сообщается с внешней средой посредством перепончатого выроста, у костистых рыб подобный вырост саккулюса заканчивается слепо.
Работами Динкграафа и Фриша было подтверждено, что слуховая функция зависит от нижней части лабиринта — саккулюса и лагены.
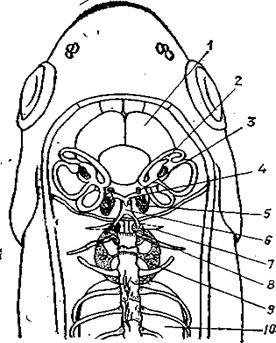
| Рис.92. Расположение лабиринта, веберова аппарата и плавательного пузыря у карпа: 1 - мозг; 2- утрикулюс; 3 -саккулюс; 4 - объединительный канал; 5 - лагена; 6 - перилимфатический проток; 7- стапес; 8- инкус; 9 - малеус; 10 -плавательный пузырь |
Лабиринт связан с плавательным пузырем цепью веберовских косточек (карповые, обыкновенные сомы, харациновые, гимнотовые), и рыбы способны воспринимать высокие звуковые тона. С помощью плавательного пузыря происходит трансформация звуков высокой частоты в низкочастотные колебания (смещения), которые воспринимаются рецепторными клетками. У некоторых рыб, не имеющих плавательного пузыря, эту функцию выполняют воздушные полости, связанные с внутренним ухом.
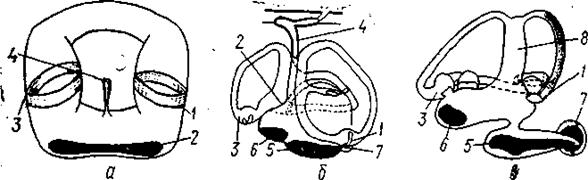
Рис.93. Внутреннее ухо или лабиринт рыб:
а— миксины; б — акулы; в — костистые рыбы;
1 - задняя криста; 2-криста горизонтального канала; 3- передняя криста;
4-эндолимфатический проток; 5- макула саккулюса, 6- макула утрикулюса; 7 - макула лагены; 8 - общая ножка полукружных каналов
Рыбы обладают также удивительным «прибором» - анализатором сигналов. Благодаря этому органу рыбы из всего хаоса окружающих их звуков и колебательных проявлений способны выделить нужные и важные для них сигналы, даже такие слабые, которые находятся на стадии возникновения или на грани затухания.

Рыбы способны усиливать эти слабые сигналы и затем воспринимать их анализирующими образованиями.
Плавательный пузырь, как полагают, играет роль резонатора и преобразователя звуковых волн, который увеличивает остроту слуха. Он выполняет также звукообразовательную функцию. Рыбы широко пользуются звуковой сигнализацией, они способны и воспринимать, и издавать звуки в широком диапазоне частот. Рыбами хорошо воспринимаются инфразвуковые колебания. Частоты равные 4-6 герцам действуют губительно на живые организмы, так как эти колебания входят в резонанс с колебаниями самого тела или отдельных органов и разрушают их. Не исключено, что рыбы реагируют на приближение ненастной погоды благодаря восприятию низкочастотных акустических колебаний, исходящих от надвигающихся циклонов.
Рыбы способны «предсказывать» изменения погоды задолго до их наступления, рыбы эти изменения фиксируют по разнице силы звуков, а возможно, и по уровню помех для прохождения волн определенного диапазона.
12.3Механизм равновесия тела у рыб. У костистых рыб утрикулюс является основным рецептором положения тела. Отолиты с помощью студенистой массы соединены с волосками чувствительного эпителия. При положении головы теменем вверх отолиты давят на волоски, при положении теменем вниз висят на волосках, при боковых положениях головы отмечается разная степень натяжения волосков. С помощью отолитов рыба принимает правильное положение головы (теменем вверх), следовательно, и туловища (спиной вверх). Для сохранения правильного положения тела также имеет значение информация, поступающая от зрительных анализаторов.
Фриш установил, что при удалении верхней части лабиринта (утрикулюса и полукружных каналов) у гольянов нарушается равновесие, рыбы ложатся на бок, брюшко или на спину на дне аквариума. При плавании они также принимают разное положение тела. Зрячие рыбы быстро восстанавливают правильное положение, а слепые равновесие восстановить не могут. Таким образом, полукружные каналы имеют большое значение в поддержании равновесия, кроме того, с помощью этих каналов происходит восприятие изменения скорости движения или вращения.
В начале движения или при его ускорении эндолимфа несколько отстает от движения головы и волоски чувствительных клеток отклоняются в сторону, противоположную, движению. При этом раздражаются окончания вестибулярного нерва. При остановке или замедлении движения эндолимфа полукружных каналов по инерции продолжает двигаться и отклоняет волоски чувствительных клеток по ходу движения.
Изучение функционального значения различных отделов лабиринта для восприятия звуковых колебаний проводилось с помощью исследования поведения рыб на основе выработки условных рефлексов, а также с помощью электрофизиологических методов.
В 1910 г. Пипер обнаружил появление токов действия при раздражении нижней частей лабиринта — саккулюса свежеубитой рыбы и отсутствие таковых при раздражении утрикулюса и полукружных каналов.
Позднее Фролов экспериментально подтвердил восприятие звуковых колебаний рыбами, проводя эксперименты на треске, применяя условно-рефлекторную методику. Фриш вырабатывал условные рефлексы на свист у карликовых сомиков. Штеттеэ. у сомов, гольянов и гольцов вырабатывал условные рефлексы на определенные звуки, подкрепляя их дачей мясных крошек, а также вызывал торможение пищевой реакции на другие звуки, ударяя рыбу стеклянной палочкой.
Органы локационной чувствительности рыб. Способности рыб к эхолокации, осуществляется не органами слуха, а самостоятельным органом - локационного органа чувств. Эхолокация относитсяковторому типу слуха. В боковой линии рыб имеется радар и сонар - составляющие органа локации.
Рыбы для своей жизнедеятельности используют электролокацию, эхолокацию, даже термолокацию. Электролокацию, часто называют шестым органом чувств рыб. Элекролокация хорошо развита у дельфинов и летучих мышей. Эти животные используют ультразвуковые импульсы частотой 60000-100000 герц, длительность посылаемого сигнала равна 0,0001 секунды, интервал между импульсами составляет 0,02 секунды. Это время требуется для анализа головным мозгом полученной информации и формирование специфической ответной реакции организма. У рыб это время немного короче. При электролокации, где скорость посылаемого сигнала равна 300000 км/с, времени для анализа отраженного сигнала у животного нет, посланный сигнал отразится и будет воспринят практически в одно и то же время.
Пресноводные рыбы не могут использовать для локации ультразвук. Для этого рыбы должны постоянно двигатся, а рыбам значительное время необходимо отдыхать. Дельфины же круглые сутки находятся в движении, у них попеременно отдыхает то левая, то правая половина мозга. Рыбы для локации используют низкочастотные волны широкого диапазона. Считается, что эти волны служат рыбам для коммуникационных целей.
Гидроакустические исследования показали, что рыбы слишком "болтливы" для неразумного существа, слишком много звуков они производят, притом "разговоры" ведутся на частотах, находящихся за пределами нормального восприятия их основным органом слуха, т.е. их сигналы более целесообразны в качестве посылаемых радарами рыб локационных сигналов. Низкочастотные волны плохо отражаются от мелких предметов, меньше поглощаются водой, слышны на большие расстояния, распространяются равномерно во все стороны от источника звука, их использование для локации дает рыбам возможность панорамного "видения - слышания" окружающего пространства.
12.5 ХЕМОРЕЦЕПЦИЯ Взаимоотношения рыб с внешней средой объединяют в две группы факторов: абиотические и биотические. Физические и химические свойства воды, действующие на рыб, называют абиотическими факторами.
Восприятие животными химические вещества с помощью рецепторов - одна из форм реакции организмов на воздействие внешней среды. У водных животных специализированные рецепторы контактируют с веществами, находящимися в растворенном состоянии, поэтому характерное для наземных животных четкое разделение на рецепторы обоняния, воспринимающие летучие вещества, и рецепторы вкуса, воспринимающие вещества, находящиеся в твердом и жидком состоянии, у водных животных не проявляется. Однако морфологически и функционально органы обоняния у рыб достаточно хорошо обособлены. Основываясь на отсутствии специфичности в функционировании, локализации и связи с нервными центрами, принято объединять вкус и общее химическое чувство понятием «химический анализатор», или «необонятельная хеморецепция».
ОРГАН ОБОНЯНИЯ относится к группе химических рецепторов. Органы обоняния рыб расположены в ноздрях, расположеных впереди каждого глаза, форма и величина которых меняется в зависимости от экологии. Представляют собой простые ямки со слизистой оболочкой, пронизанные разветвлением нервов, ведущие в слепой мешок с чувствительными клетками идущих от обонятельной доли головного мозга.
У большинства рыб каждая из ноздрей поделена перегородкой на автономные передние и задние носовые отверстия. В некоторых случаях, носовые отверстия одинарные. В онтогенезе носовые отверстия у всех рыб первоначально одинарны, т.е. не поделены перегородкой на переднюю и заднюю ноздри, которые обособляются лишь на более поздних стадиях развития.
Расположение ноздрей у различных видов рыб зависит от их образа жизни и развития других органов чувств. У рыб с хорошо развитым зрением носовые отверстия расположены на верхней стороне головы между глазом и концом рыла. У селяхше ноздри располагаются на нижней стороне и приближены к ротовому отверстию.
Относительная величина ноздрей тесно связана со скоростью движения рыбы. У рыб, плавающих медленно, ноздри сравнительно больше, причем перегородка между передним и задним носовыми отверстиями имеет вид вертикально расположенного щитка, направляющего воду обонятельную капсулу. У рыб быстрых, носовые отверстия чрезвычайно маленькие, так как при больших скоростях встречного обтекающего конька, вода в носовой капсуле смываются достаточно быстро, через относительно маленькие отверстия передних ноздрей. У придонных рыб, у которых роль обоняния в общей системе рецепции очень существенна, передние носовые отверстия вытягиваются в виде трубочек и приближаются к ротовой щели или даже свешиваются с верхней челюсти в низ, это имеет место, у Typhleotris, Anguilla, Mnreana и др.
Пахучие вещества, растворенные в воде, попадают на слизистую оболочку обонятельной области, раздражают окончания обонятельных нервов, отсюда сигналы поступают в головной мозг.
 Посредством обоняния рыбы получают информацию об изменениях внешней среды, различают пищу, находят свою стаю, партнеров во время нереста, обнаруживают хищников, вычисляют добычу. На коже у некоторых видов рыб расположены клетки, которые при ранении кожи выделяют в воду «вещество страха», которое является для других рыб сигналом об опасности. Рыбы активно используют химическую информацию для подачи сигналов тревоги, предупреждения об опасности, привлечения особей противоположного пола. Особенно важен этот орган для рыб, обитающих в мутной воде, где наряду с осязательной и звуковой информацией рыбы активно используют и обонятельную систему. Обоняние оказывает большое влияние и на работу многих органов и систем организма, тонизируя или угнетая их. Известны группы веществ, положительно (аттраканты) или отрицательно (репелленты) действующие на рыб. Обоняние тесно связано с другими органами чувств: вкуса, зрения и равновесия.
Посредством обоняния рыбы получают информацию об изменениях внешней среды, различают пищу, находят свою стаю, партнеров во время нереста, обнаруживают хищников, вычисляют добычу. На коже у некоторых видов рыб расположены клетки, которые при ранении кожи выделяют в воду «вещество страха», которое является для других рыб сигналом об опасности. Рыбы активно используют химическую информацию для подачи сигналов тревоги, предупреждения об опасности, привлечения особей противоположного пола. Особенно важен этот орган для рыб, обитающих в мутной воде, где наряду с осязательной и звуковой информацией рыбы активно используют и обонятельную систему. Обоняние оказывает большое влияние и на работу многих органов и систем организма, тонизируя или угнетая их. Известны группы веществ, положительно (аттраканты) или отрицательно (репелленты) действующие на рыб. Обоняние тесно связано с другими органами чувств: вкуса, зрения и равновесия.
В различные времена года обонятельные ощущения у рыб не одинаковы, они обостряются весной и летом, особенно в теплую погоду.
 Рыбы, ведущие ночной образ жизни (угорь, налим, сом) обладают высокоразвитым обонянием. Обонятельные клетки этих рыб способны реагировать сотые доли концентраций аттракантов и репеллентов.
Рыбы, ведущие ночной образ жизни (угорь, налим, сом) обладают высокоразвитым обонянием. Обонятельные клетки этих рыб способны реагировать сотые доли концентраций аттракантов и репеллентов.
 Рыбы способны почувствовать разведение в пропорции один к миллиарду экстракта из мотыля, карась чувствует аналогичную концентрацию нитробензола, более высокие концентрации менее привлекательны для рыб. Для обонятельного эпителия стимуляторами служат аминокислоты, некоторые из них или же их смеси имеют сигнальное значение для рыб. Например, угорь находит моллюска по выделяемому им комплексу, состоящему из 7 аминокислот. Позвоночные животные ориентируются на смесь основных запахов: мускусный, камфорный, мятный, эфирный, цветочный, острый и гнилой.
Рыбы способны почувствовать разведение в пропорции один к миллиарду экстракта из мотыля, карась чувствует аналогичную концентрацию нитробензола, более высокие концентрации менее привлекательны для рыб. Для обонятельного эпителия стимуляторами служат аминокислоты, некоторые из них или же их смеси имеют сигнальное значение для рыб. Например, угорь находит моллюска по выделяемому им комплексу, состоящему из 7 аминокислот. Позвоночные животные ориентируются на смесь основных запахов: мускусный, камфорный, мятный, эфирный, цветочный, острый и гнилой.
Рецепторы обоняния у рыб, как и у прочих позвоночных, парные и расположены на передней части головы. Лишь у крутлоротых непарные. Обонятельные рецепторы находятся на слепом углублении - ноздре, дно которой выстлано обонятельным эпителием, располагающимся на поверхности складок. Складки, расходясь радиально от центра, образуют обонятельную розетку.
У разных рыб обонятельные клетки расположены на складках по-разному: сплошным слоем, разреженно, на гребнях или в углублении. Ток воды, несущий молекулы пахучих веществ, попадает к рецептору через переднее отверстие, часто отделеное от выходного заднего отверстия лишь складкой кожи. Однако у некоторых рыб входное и выходное отверстия заметно разделены и далеко стоят друг от друга. Передние (входные) отверстия у ряда рыб (угорь, налим) находятся близко к концу рыла и снабжены кожными трубочками. Считается, что этот признак свидетельствует о значительной роли обоняния в поисках пищевых объектов. Движение воды в обонятельной ямке может создаваться либо движением ресничек на поверхности выстилки, либо сокращением и расслаблением стенок специальных полостей - ампул, либо в результате движения самой рыбы.
Рецепторные обонятельные клетки, имеющие биполярную форму, относятся к категории первичных рецепторов, т. е. сами регенерируют импульсы, содержащие информацию о раздражителе и передают их по отросткам к нервным центрам. Периферический отросток обонятельных клеток направляется к поверхности рецепторного слоя и заканчивается расширением - булавой, на апикальном конце которой имеется пучок волосков или микроворсинок. Волоски пронизывают слой слизи на поверхности эпителия и способны к движениям.
Обонятельные клетки окружены опорными клетками, которые содержат овальные ядра и многочисленные гранулы разного размера. Здесь же расположены базальные клетки, не содержащие секреторных гранул. Центральные отростки рецепторных клеток, не имеющие миелиновой оболочки, пройдя базальную мембрану эпителия, образуют пучки до нескольких сотен волокон, окруженные мезаксоном шванновской клетки, причем тело одной клетки может охватывать много пучков. Пучки сливаются в стволики, образующие обонятельный нерв, соединяющийся с обонятельной луковицей.
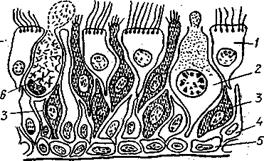
| Рис. 95. Строение обонятельной выстилки:. 1-ресничная клетка; 2- грушевидная секреторная клетка; 3-обонятельные луковицы; 4-опорные клетки; 5- базальные клетки; 6-бокаловидная секреторная клетка |
Строение обонятеельной выстилки у всех позвоночных сходно (рис. 95), что свидетельствует о сходстве механизма контактной рецепции. Однако сам этот механизм еше не вполне ясен. Одна из них связывает возможности распознавания запахов, т. е. молекул пахучих веществ, с избирательной специфичностью индивидуальных рецепторов к запахам. Такова стереохимическая гипотеза Эймура. согласно которой, имеется семь типов активных мест на обонятельных клетках, а молекулы веществ обладающих сходными запахами, имеют одинаковую форму активных частей, которые подходят к активным точкам рецептора, как " ключ" к замку. Другие гипотезы связывают возможность различения запахов с различиями в распределении веществ, адсорбируемых слизью обонятельной выстилки, по ее поверхности. Ряд исследователей полагают, что распознавание запахов обеспечивают эти два механизма, дополняя друг друга.
Ведущая роль - в обонятельной рецепции принадлежат, волоскам и булаве обонятельной клетки, которые обеспечивают специфическое взаимодействие молекул одорантов с мембраной клетки и перевод эффекта взаимодействия в форму электрического потенциала. Как уже было сказано, аксоны обонятельных рецепторных клеток образуют обонятельный нерв, входящий в обонятельную луковицу, являющуюся первичным центром обонятельного рецептора.
Обонятельная луковица относится, как считает А. А. Заварзин, к экранным структурам. Для нее характерно расположение элементов в виде последовательных слоев, причем нервные элементы связаны между собой не только в пределах слоя, но и между слоями. Таких слоев выделяют обычно три: слой обонятельных клубочков с межклубочковыми клетками, слой вторичных нейронов с митральными и кисточковыми клетками, гранулярный слой.
Информацию в высшие обонятельные центры у рыб передают вторичные нейроны и клетки гранулярного слоя. Наружная часть обонятельной луковицы состоит из волокон обонятельного нерва, контакт которых с дендритами вторичных нейронов происходит в обонятельных клубочках, где наблюдается ветвление тех и других окончаний. В одном обонятельном клубочке сходятся несколько сотен волокон обонятельного нерва. Слои обонятельной луковицы обычно расположены концентрически, но у некоторых видов рыб (щука), они лежат последовательно в рострокаудальном направлении.
Обонятельные луковицы рыб анатомически хорошо обособлены и бывают двух типов: сидячие, прилегающие к переднему мозгу; стебельчатые, расположенные сразу за рецепторами (очень короткие обонятельные нервы).
У тресковых, обонятельные луковицы связаны с передним мозгом длинными обонятельными трактами, которые представлены медиальным и латеральным пучками, оканчивающимися в ядрах переднего мозга.
Обоняние как способ получения информации об окружающем мире имеет для рыб весьма существенное значение. По степени развития обоняния рыб, как и других животных, обычно делят на макросматиков и микросматиков. Это деление связанно с различной широтой спектра воспринимаемых запахов.
У макресматиков органы обоняния способны воспринимать большое количество различных запахов, т. е. обоняние используется ими в более разнообразных ситуациях.
Микросматики воспринимают обычно неболышое количество запахов - в основном особей своего вида и половых партнерш. Типичным представителем макросматиков являётся обыкновенний угорь, микросматиков - щука, трехиглая колюшка. Для восприятия запаха иногда, видимо, достаточно попадания на обонятельный рецептор, нескольких молекул вещества.
Обоняние может играть руководящую роль в поисках пищи, особенно у ночных и сумеречных хищников, таких, как угорь. С помощью обоняния рыбы могут воспринимать партнеров по стае, находить особей другого пола в период размножения. Например, гольян может различать среди особей своего вида партнера. Рыбы одного вида способны воспринимать химические соединения, выделяемые кожей других рыб при ранении.
Изучение миграций проходных лососей показало, что на этапе входа в нерестовые реки они отыскивают именно ту реку, где сами вывелись из икры, ориентируясь по запаху воды, запечатленному в памяти на этапе молоди (рис.96). Источниками запаха, видимо, являются виды рыб, постоянно обитающие в реке. Эта способность была использована для направления мигрирующих производителей в определенный участок. Молодь кижуча выдерживалась в растворе морфолина с концентрацией 0~5 М, а затем, после их возвращения в период нереста в родную реку, привлекалась тем же раствором к определенному месту в водоеме.
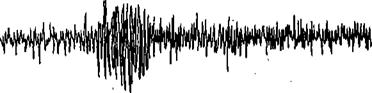

Рис. 96. Биотоки обонятельного мозга лосося при орошении обонятельных ямок; 1, 2 - дистиллированной водой; 3 - водой из родной реки; 4, 5, 6 - водой из чужих озер.
Рыба обладает обонянием, которое сильнее развито у нехищных рыб. Щука, например, при поисках пищи не пользуется обонянием. При ее быстром броске за добычей обоняние не может играть существенной роли. Другой хищник - окунь при передвижениях в поисках пищи обычно плавает тихо, подбирая со дна всевозможные личинки, он обонянием в данном случае пользуется как органом, наводящим на пищу.
Орган вкуса имеется почти у всех рыб, вкусовые ощущения передаются большинству из них через губы и рот. Поэтому рыба не всегда проглатывает схваченный корм, особенно если он не пришелся ей по вкусу.
Вкус - это ощущение, возникающее при действии на орган вкуса пищевых и некоторых не пищевых веществ. Орган вкуса тесно связан с органом обоняния и относится к группе химических рецепторов. Вкусовые ощущения у рыб появляются при раздражении чувствительных, осязательных клеток - вкусовых сосочек или так называемых вкусовых почек, луковиц, расположенных в ротовой полости в виде микроскопических вкусовых клеток, на усиках, по всей поверхности тела, особенно на кожных выростах. (Рис.97)
Главными восприятиями вкусовых ощущении являются четыре составляющих: кислое, сладкое, соленое и горькое. Остальные виды вкуса представляют собой комбинации этих четырех ощущений, причем вкусовые ощущения у рыб могут вызывать только вещества, растворенные в воде.
Минимально ощутимая разница в концентрации растворов веществ порог различия - постепенно ухудшается при переходе от слабых к более сильным концентрациям. К примеру, однопроцентный раствор сахара обладает практически максимально сладким вкусом, и дальнейшее увеличение его концентрации не изменяет вкусового ощущения.
Появление вкусовых ощущений может быть вызвано действием на рецептор неадекватных раздражителей, например, постоянного электрического тока. При длительном соприкосновении какого-либо вещества с органом вкуса постепенно притупляется его восприятие, в конце концов, это вещество покажется рыбе совершенно безвкусным происходит адаптация.
Вкусовой анализатор также может влиять на некоторые реакции организма, на деятельность внутренних органов. Установлено, что рыбы реагируют практически на все имеющие вкус вещества и обладают при этом поразительно тонким вкусом. Положительные или отрицательные реакции рыб определяются их образом жизни и, прежде всего, характером их питания. Положительные реакции на сахар свойственны животным, питающимся растительной и смешанной пищей. Ощущение горечи у большинства живых существ вызывает отрицательную реакцию, но не у тех, которые питаются насекомыми.
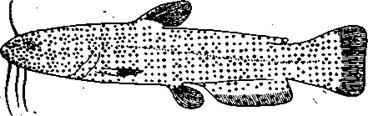
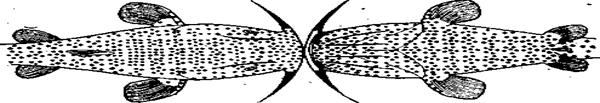
Рис.97. Расположение вкусовых почек на теле сома показано точками. Каждая точка означает 100 вкусовых почек
Механизм вкусовой рецепции. Четыре основных вкусовых ощущения - сладкое, горькое, кислое и соленое - воспринимаются благодаря взаимодействию молекул вкусовых веществ с четырьмя молекулами белков. Комбинации этих типов и создают конкретные вкусовые ощущения. У большинства рыб вкус играет роль контактной рецепции, поскольку пороги вкусовой чувствительности сравнительно высоки. Но у некоторых рыб вкус может приобретать функции дистантного рецептора. Так, пресноводный сомик с помощью вкусовых рецепторов способен локализовать пищу на расстоянии около 30 длин тела. При отключении вкусовых рецепторов эта способность исчезает. С помощью общей химической чувствительность рыбы способны улавливать изменения солености до 0,3% концентрации отдельных солей, изменения концентрации растворов органических кислот (лимонная) до 0,0025 М (0,3 г/л), изменения рН порядка 0,05—0,07 концентрации углекислоты до 0,6 г/л.
Необонятельная хеморецепция у рыб осуществляется вкусовыми почками и свободными окончаниями блуждающего, тройничного и некоторых спинномозговых нервов. Структура вкусовых почек сходна у всех классов позвоночных. У рыб они обычно имеют овальную форму и состоят из 30—50 вытянутых клеток, апикальные концы которых образуют канал. К основанию клеток подходят окончания нерва. Это типичные вторичные рецепторы. Они располагаются в ротовой полости, на губах, жабрах, в глотке, на коже головы и туловища, на усиках и плавниках. Их количество варьирует от 50 до сотен тысяч и зависит, как и их расположение, в большей мере от экологии, нежели от вида. Размеры, количество и распределение вкусовых почек характеризует степень развития вкусовой рецепции конкретного вида рыб. Вкусовые почки передней части рта и кожи инервируются волокнами возвратной ветви лицевого нерва, а слизистой рта и жабр - волокнами языкоглоточиого и блуждающего нервов. В иннервации вкусовых рецепторов, участвуют также тройничный и смешанные нервы.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 6114; Нарушение авторских прав?; Мы поможем в написании вашей работы!