КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физиология центральной нервной системы 2 страница
|
|
|
|
Верхнечелюстная (r. maxillaris) и нижнеечелюстная (r. manolibularis) ветви, начинаются общим стволом и обособляются позади глаза, они иннервируют мышцы челюстной дуги, зубы, слизистую оболочку ротовой полости. По функции, тройничный нерв смешанный, он имеет чисто чувствующую ветвь-глазничную и смешанные (чувствующие и двигательные) верхнечелюстную и нижнечелюстную.
Лицевой нерв (n. facialis). Отходит сразу же за тройничным. Этот нерв также распадается на несколько ветвей наиболее крупные из них - глазничная, щечная, небная и подъязычная ветви.
Глазничная ветвь (r. ophtalmicus) идет вместе с одноименной ветвью тройничного нерва. Позади глазничной ветви отходит крупный ствол щечная ветвь (r. buccalis). Она в виде широкого тяжа отходит по дну глазницы, иннервируя кожные органы чувств брюшной поверхности головы. Далее общим стволом отходят две ветви: небная и подъязычная.
Небная ветвь (r. palatinus) сразу же после обособления уходит в дно черепной коробки и затем ветвится в слизистой оболочке ротовой полости.
Подъязычная ветвь (r. hyamanolibularis) проходит позади брызгальца и иннервирует мускулатуру подъязычной дуги и кожные органы чувств боков головы. Лицевой нерв, как и тройничный смешанный, его глазничная, щечная и небная ветви чувствующие, а подъязычная смешанная (чувствцющая и двигательная).
Слуховой (n. acusticus). Идет от боковой поверхности продолговатого мозга и почти сразу же входит в стенку черепной коробки, иннервируя внутреннее ухо. Это чисто чувствующий нерв.
Языкоглоточный нерв (n. glozsopharynqeus). Подходит к первой жаберной щели и здесь делится на две ветви иннервирующие соответственно ее переднюю и заднюю поверхности. Задняя ветвь иннервирует всю мускулатуру первой жаберной дуги. Этот нерв по своей функции смешанный, он имеет и чувствующие и двигательные волокна.
Блуждающий нерв (n. vagus). Отходит от задне - боковой поверхности продолговатого мозга несколькими корешками почти сразу же сливающимися в толстый нервный тяж. Иннервирует обширную область тела, его основные ветви - четыре жаберные, внутренностная и боковая.
Четыре жаберные ветви (r. branchialis) иннервируют жаберные щели от второй до пятой. Каждая ветвь пройдя к жаберной щели, делится на две передне - и заднежаберную. Переднежаберная ветвь чувствующая и заднежаберная смешанная вместе иннервируют всю мускулатуру, соответствующей жаберной дуги (от второй до пятой).
Внутренностная ветвь (r. splanchnicus) идет как продолжение основного ствола блуждающего нерва. Она входит в брюшную полость, иннервируя внутренние органы. Поверхностно расположена боковая ветвь (r. lateralis) иннервирующая органы боковой линии туловища и хвоста. Блуждающий нерв смешанный (включает и чувствующие, и двигательные волокна). У высших позвоночных животных обособляются еще две пары головных нервов.
Подъязычный нерв (n. hypoqlozzus) впервые четко обособляется у пресмыкающихся. Зачатки этих нервов есть у акуловых рыб в виде нервных корешков, отходящих позади блуждающего нерва в области затылочной стенки мозгового черепа.
Спинномозговые нервы (n. spinalis) посегментально отходят от спинного мозга (medulla spinalis). Иннервирует, соответствующие сегменты тела и по своей функции представляют собой смешанные нервы, состоящие из чувствующих и двигательных волокон.
Органы боковой линии. Органы боковой линии представляют собой сформировавшуюся в процессе эволюции высокоспециализированную сенсорную систему, существующую у первичноводных позвоночных животных, круглоротых, рыб. Органы боковой линии расположены в коже и подкожных структурах, однако по своему строению они сходны с рецепторами внутреннего уха. В эмбриональный период оба этих типа рецепторов развиваются из одного и того же эктодермального утолщения височной, области, т.е. имеют общее происхождение, орган боковой линии выполняет четко выраженную механорецепторную функцию.
Адекватным стимулом для этой системы являются смещения водной среды, возникающие при различных колебаниях водной среды, перепадах давления в потоках воды, в том числе и при действии низко частичных звуковых волн.
В составе боковой линии некоторых рыб наряду с механорецепторными структурами существуют и нижесенсорные образования, предназначенные для восприятия слабых электрических полей водной среды. Для этого чтобы различать эти два типа сенсорных приборов, механорецепторные органы стали называть обычными, а электрорецепторные образования - специализированными.
Источниками сигналов для электроценторов могут служить самые разнообразные электрические явления, возникающие в водной среде в результате различных физических и физико - химических процессов, а также биологическими процессами, связанными с жизнедеятельностью животных.
Всех рыб, обладающих развитым электрорецепторным аппаратом, в соответствии с их способностью к генерации электрических полей принято подразделять на 3 группы;
1) группу составляют неэлектрические рыбы, не имеющие электрических органов. К ним относятся большинство пластиножаберных, за исключением семейства обыкновенных скатов, а также ряд представителей отряда сомовых, например карликовый сомик, туркестанский сомик.
2) группу составляют слабоэлектрические рыбы, их электрические органы генерируют разряды небольшой величины, не превышающие по амплитуде 10-15В. В эту группу входят, представители подотряда электрических угрей, обитающих в реках Южной Америки, представители отряда мормирообразных, включающих два подотряда – гимнархид и мормирид, живущих в африканских реках, и представители семейства обыкновенных скатов.
3) К группе относятся сильноэлектрические рыбы, способные генерировать как низковольтные, так и высокоамплитудные разряды, достигающие нескольких сотен вольт.
Механорецепторные структуры боковой линии. Основу органов боковой линии представляют специализированные волосковые клетки — нейромасты (рис.), располагающиеся на основной мембране. Каждый нейромаст имеет различные виды волосков: один крупный — киноцилий и пучок из 40— 50 мелких — стереоцилий. Волоски погружены в желеобразную среду — купулу. Несколько нейромастов объединяются и образуют чувствительный бугорок. Кроме того, имеются опорные эпителиальные клетки.
Наиболее просто органы боковой линии устроены у круглоротых и некоторых костистых рыб (колюшка), у которых они представлены продольными рядами чувствительных клеток, расположенных на поверхности тела,— поверхностные органы. У акул и химер нейромасты расположены на дне желобка, проходящего по бокам тела, у ганоидов и большинства костистых рыб эти клетки располагаются на дне канала, сообщающегося с поверхностью тела небольшими тонкими ходами. Это канальные органы.
У многих рыб органы боковой линии располагаются не только по бокам тела, но и в области головы, на жаберных крышках и нижней челюсти. Степень развития боковой, линии у разных видов рыб различна. У верховки и корюшки она распространяется на небольшое расстояние позади головы, у других рыб она хорошо развита на всем протяжении туловища.
У костистых рыб и амфиогии одиночный эпидермальный орган, или невромаст состоит из 20-60 рецепторных клеток, снабженных усилиями или волосками окружающих их опорных клеток и желатинообразной купулы, покрывающей сверху волосковые клетки.Совокупность рецепторных и опорных клеток образует сенсорный эпителий. Волосковая клетка имеет длину в среднем 30-40 и ширину 7-10 мкм. Ее цилиарный аппарат состоит из нескольких десятков стереоцилии и одной киноцилии, длинной около 40 мкм. Стереоцими и киноцилия своими дистальными концами погружены в капсулу, которая у свободных невромастов может выступать над поверхностью кожи до 100 мкм.
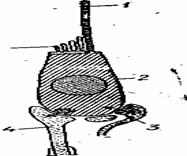
| Рис.80. Строение нейромастов - волосковой клетки с афферентной и эфферентной иннервацией: 1- киноцилий; 2 - волосковая клетка; 3 - тормозящий нерв; 4-афферентный (чувствительный) нерв; 5 - стереоцилий |
Вследствие того, что киноцилия располагается всегда асимметрично относительно пучка стереоцилии, рецепторная клетка оказывается морфологически поляризованной. У рыб одиночная волосковая клетка иннервируется обычно несколькими первичными волосками, образующими на базальной части клетки окончания двух типов: гранулированные и негранулированные органы боковой линии иннервируются передним и задним нервами боковой линии. Передний нерв снабжает рецепторные структуры на голове, а задний на туловище животного. Нервы боковой линии представляют собой периферические отростки ганглиозных клеток, лежащие внутрикраниально. Передний нерв боковой линии образован ветвью лицевого или тройничного нервов, а задний ветвями блуждающего и языкоглоточного нервов.
Механорецепторные структуры боковой линии представлены двумя разновидностями чувствительных образований, свободными невромастами и канальными органами. Основное отличие между ними заключается в особенностях структуры вспомогательного аппарата, благодаря которому свободные невроматы непосредственно контактируют с наружной средой, а канальные органы такого контакта не имеют и внешний стимул вызывает, прежде всего, перемещение канальной жидкости, которое и воспринимается рецепторами. У костистых рыб сенсорные каналы состоит из 5 основных ветвей: главного латерального канала, надвисочного, височного, над и подглазничного каналов.
Функциональные свойства электроцепторной структуры.
Локальные электрические явления в рецепторах. В рецепторах боковой линии, как и в других вторичночувствующих рецепторах, функцию генераторного потенциала выполняет возбуждающий постсинаптический потенциал (ВПСП). Частое ограничение системы осуществляется не на уровне рецепторов, а за счет механических свойств окружающих их элементов, взаимодействующих с водной средой.
Синаптическая передача. Между волосковой клеткой и окончаниями сенсорного нейрона имеются синоптические контакты, характерные для синапсов с химическим механизмом передачи.
Эфферентная регуляция. Эфферентные влияния на деятельность механорецепторов боковой линии носят тормозной характер и воздействуют на спонтанную и вызванную активность рецепторов.
Эфферентные влияния, снижая общий уровень фоновой активности рецепторов, уменьшают не только фоновый шум, существующий в данной сенсорной системе, но и величину рецептивного поля, что имеет важное значение в локализации источника стимула в пространстве.Эфферентные импульсы в органах боковой линии рыб могут быть вызваны афферентными разрядами, причем таким действием обладают также и контралатеральные афферентные входы. В эфферентной системе в качестве медиатора выделяется ацетилхолин.
Импульсивная активность. В состоянии покоя в нервных волокнах, иннервирующих невромасты бок линии, регистрируется довольно высокий уровень фоновой активности, достигающий 100 имп./сек. наличие фоновой активности в нервных волокнах отражает фоновую активность рецепторов и связано со спонтанным выделением медиатора из волосковых клеток.
Специализированные рецепторы боковой линии отличаются большим разнообразием. Наиболее удачная – это классификация Сабо, согласно которой все электрорецепторы следует подразделять на ампулированные и бугорковые. В ампулированных органах рецепторные клетки сообщают с наружной средой при помощи специального канала, тогда как бугорковые органы такого канала не имеют. Ампулированные рецепторы имеются у всех морских и пресноводных рыб, обладающих электроцепторным аппаратом. Бугорковые рецепторы встречаются только у слабоэлектрических рыб.
Ампулированные рецепторы. К ним относятся не имеющие специального наименования ампулированные органы слабоэлектрических рыб, ампулы Лоренцини пластиножаберных рыб и малые ямковые органы сомовых рыб. Отдельный орган состоит из канала, идущего от поверхности кожи и заканчивающегося слепым расширением - ампулой, в основании которой располагается сенсорный эпителии. У слабоэлектрических рыб длина канала варьирует от 100 до 200 мкм.
Ампулярные органы распределены по всей поверхности тела, но их концентрация в области головы особенно велика. Иначе организованы ампулы Лоренцини, встречающиеся у пластиножаберных рыб и единственной костистой рыбы Plotosus. Длина каналов у этих рецепторов в зависимости от среды обитания варьирует от 500 мкм. У пресноводного ската Potamotrygon до нескольких десятков сантиметров у крупных морских скатов.
Каналы расположены под кожей и имеют различную длину, так как берут начало в различных участках тела и сходятся к капсулам, где заканчиваются ампулярным расширением, на дне которого расположены чувствительные клетки. На одной рецепторной клетке может быть несколько синапсов, а одно и то же нервное окончание может контактировать сразу с двумя соседними рецепторными клетками.
Ампулярные каналы заполнены желеобразным веществом, состоящим главным образом из мукополисахаридов. Рецепторы апмулированных образований функцианируют в условиях окружения некоторой специфической среды, отличающейся повышенным содержанием ионов калия по сравнению с наружной средой и жидкостями тела. Особенности ампулярного канала придают ему свойства идеального морского кабеля, обеспечивающего прохождение сигнала вдоль него почти без ослабления.
Бугорковые рецепторы располагаются в инвагинациях эпидермальной мембраны. Отдельные органы содержат рецепторные клетки, находящиеся в эпидермальной капсуле, не имеющей прямого контакта с наружной средой количество рецепторных клеток в капсуле варьирует от 1 до 35 у мормирид и от 10 до 100 у гипнотид.
Функциальные свойства. Ампулированные рецепторы разных типов имеют принципиально сходное строение, характеризующееся наличием ампулоподобного органа в качестве основной структурной единицы.
Ампулированные рецепторы слабоэлектрических костистых рыб менее чувствительны к электрическим полям. Исследования электрической чувствительности бугорковых рецепторов слабоэлектрических рыб показывают, что этот класс рецепторов оказывается менее чувствительным к электрической стимуляции, чем ампулированные рецепторы.
Ампулированные рецепторы наряду с высокой электрической чувствительностью обладают сильно развитой чувствительностью к изменению температуры окружающей среды. Снижение температуры воды вызывает временное повышение частоты импульсации, а при повышении температуры наблюдаются противоположные изменения активности.
Термочувствительной является область сенсорного эпителия. Механическая чувствительность ампулированных рецепторов выражена слабо. В отношении химической чувствительности рецепторов ампул Лоренцини (восприятие изменении солености, содержание различных ионов, углекислого газа) обусловлена возникновением соответствующих полей электрохимического происхождения.
Центральные структуры системы боковой линии:Продолговатый мозг. Нервы боковой линии оканчиваются в ядрах акустико - латеральной области (area acousticolateralis). Акустика - латеральная область, продолговатого мозга является неоднородным образованием и состоит из нескольких ядер. У гонадно - и поперечноротых рыб ее принято подразделять на дорсальное, медиальное и вентральное ядра. Вентральное ядро связано со слуховой системой. Медиальное ядро, образованная клетками дорсолатеральной части продолговатого мозга, получает волокна, как от переднего, так и от заднего нервов боковой линии.
Мозжечок рыб принято подразделять на тело, вальвулу и вестибулолатеральную долю. Последняя представляет собой наиболее каудальную часть мозжечка, которая является непосредственным продолжением акустико- латереальной области. Lobus vestibulolateralis разована плотной массой гранулярных клеток вдоль латерального края доли и auriculi -другой группой гранулярных клеток, расположенных более медиально. Развитие мозжечка связано с общими специализированными электрорецепторными образованиями на теле животного.
Средний и передний мозг. Информация от органов боковой линии поступает в структуры среднего мозга. Соответствующий центр расположен в torussemicircularis покрышки среднего мозга. Это ядро может быть подразделено на две части: pars lat. u parsmed. Nucl. Lat, которое связано с акустик – латеральной областью с помощью латерального лемнического пути, о чем свидетельствуют эксперименты с дегенерацией нервных волокон после экспериментального повреждения долей боковой линии продолговатого мозга. Латеральная система рыб широко представлена в различных отделах мозга животных.
В продолговатом мозге пластиножаберных рыб ВП локализуются в пределах акустико - латеральной области. У пластиножаберных рыб ответы регистрируются в аурикулах и задней доле тела мозжечка.
Средний Он является коллектором информации, идущей от различных сенсорных систем. У неэлектрических костистых рыб акустико- латеральная система представлена в двух ядрах среднего мозга.
Биологическое значение органов боковой линии. Боковая линия дает возможность рыбе ориентироваться, чувствовать течение воды или приближение в темноте различных предметов. Органы боковой линии расположены в канале, погруженном в кожу, который сообщается с внешней средой при помощи отверстий в чешуе. Снаружи виден ряд отверстий. Эти отверстия связаны с каналом, расположенным в коже. В канале находятся чувствительные клетки, нервные окончания соединенные с нервом, идущим под кожей. Боковая линия воспринимает направление и силу тока воды. Благодаря боковой линии даже ослепленная рыба не натыкается на препятствия и способна ловить движущуюся добычу.
Биологическое назначение механорецепторной системы боковой линии еще недостаточно выявлено. Методом выработки условных, пищевых рефлексов в сочетании с денервацией органов, удалось показать о существовании некоторых поведенческих реакций при локальном действии слабых потоков воды на голову и тело у пластиножаберных и костистых рыб, а также у амфиогии. Выявлена способность ослепленных животных воспринимать искусственные препятствия на расстоянии 0,5 км, что свидетельствует от участии органов боковой линии в ориентировочном поведении с относительно малым радиусом действия. Характерно поведение многих рыб во время нереста, когда партнеры раздражают друг друга слабыми потоками воды, возникающими при движении плавников. Отмеченные реакции наблюдаются только у животных с интактной системой боковой линии.
Одной из разновидностей механической стимуляции являются вибрационные воздействия. Частотный диапазон восприятия вибрации находится в пределах от 1- до 25 Гц. Рыбы, обитающие в поверхностных слоях воды, воспринимают вибрационные стимулы в виде волн, распространяющихся по поверхности воды. В боковой линии этих рыб имеются невромасты не только вдоль боковой поверхности, но и на нижней части туловища. Такая система организации невромастов, различно расположенных на теле рыб, способствует как дифференцированному восприятию внешних стимулов, так и появлению большей чувствительности и точности локализации источников раздражении.
Относительно роли органов боковой линии в формировании стайного поведения рыб сведения противоречивы. По данным Дислера, становление стайности наблюдается у молоди карповых и окуневых рыб в конце личиночного начало малькового периода, когда органы боковой линии уже хорошо развиты. Очень интересно устроена у рыбы так называемая боковая линия, ее можно заметить почти у всех рыб. Расположена она с боков на чешуе в виде черных точек (это отверстия в чешуе) и тянется от головы до хвоста. У некоторых видов рыб например у щуки, этот орган находится и на голове, на верхней и нижней челюстях.
Боковая линия—это своеобразная, очень чувствительная сигнальная установка, ею рыба чувствует направление течения воды и его силу, ею же по малейшим изменениям тока воды может чувствовать близость препятствий на своем пути: берега, камни, коряги, завалы, движущиеся в воде предметы, проплывающую рыбу. При помощи боковой линии хищные рыбы получают сигнал о приближении живой добычи и месте ее нахождения, а “добыча” получает в это же время противоположный сигнал надвигающейся опасности.
Боковая линия особенно хорошо развита у щуки, трудно установить, каким органом — зрением или боковой линией щука более руководствуется при поисках корма. Это необходимо учесть рыболовам-спиннингистам и изготовлять свои приманки (насадки), блесны, рассчитывая не только на восприимчивость их зрением щуки, но и колебаниями блесны, раздражителем боковой линии, и чем эти колебания будут более сходны с колебаниями от живой, плывущей рыбки, тем более вероятной будет хватка щуки.
Чувствительность бокового органа очень велика, и рыба способна воспринимать очень незначительные колебания. Интересно, что рыба очень точно находит место, от которого исходит раздражение, и безошибочно захватывает предмет, даже если он приближается с заднего конца тела рыбы. Ослепленная щука могла хорошо захватывать двигающуюся около нее рыбу, однако, если рядом с ней клалась мертвая рыба, то щука ее не захватывала. Если же мертвую рыбу двигали около щуки, она брала ее как живую. Если повредить боковой орган, то рыба в темноте будет натыкаться на предметы, находящиеся в воде.
Последними научными данными установлены многие ранее неизвестные явления в жизни рыбы, например, рыбу удается приучить к тому, что она сможет отличать квадрат от прямоугольника, овал от круга и т. п. или брать корм лишь из кормушки, окрашенной в красный цвет. Рыба (пескарь) скоро научилась добывать корм именно из кормушки красного цвета, не обращая внимания на кормушки других цветов. Этим было точно установлено, что рыба различает форму и цвета. Однако это не значит, что рыба, различая цвета, находящиеся в неподвижном состоянии, сможет различить цвета быстро движущегося предмета в воде, например цвет “Девона” — белый он или желтый.
Предполагается, что боковой линией воспринимаются низкочастотные звуковые волны от 5 до 600 герц. Рыбы способны воспринимать весь диапазон звуковых колебаний: от инфра - до ультразвуковых. Установлено, что рыбы способны уловить в 10 раз меньшее изменение частот, нежели человек, но музыкальный слух у рыб в 10 раз хуже.
Раздражителями для чувствительных клеток боковой линии являются токи воды, вибрация и перепады давления, при этом происходит движение жидкости в каналах, приводящее к смещению волосков чувствительных клеток. Это воспринимается латеральными нервами туловища и головы.
С помощью органов боковой линии рыба ориентируется в потоке воды, воспринимает движущиеся предметы, различает низкочастотные колебания. Парные органы, находящиеся в боковой линии рыб, панорамно воспринимают звуковые колебания, что дает возможность рыбам четко устанавливать направление и место источника колебания. Рыбы выделяют ближнюю и дальнюю зоны акустического поля. В ближней зоне они четко определяют местонахождение источника колебания.
Инфразвуки или вибрации — колебания с частотой до 16 Гц — не воспринимаются рыбами во взвешенном состоянии без контакта с другими предметами и грунтом. Механические раздражения, возникающие в результате гидродинамических процессов, воспринимаются органами боковой линии. Звуковые колебания воспринимаются лабиринтовым аппаратом внутреннего уха. Низкочастотные звуковые колебания воспринимаются рецепторами плавательного пузыря и боковой линии.
Рецепторы боковой линии воспринимают звуки на близком расстоянии, с помощью внутреннего уха происходит анализ звуков, доносящихся с большого расстояния.
Рыбы могут определить и направление источника звука, но точно локализовать его не могут. Правильную локализацию источника звука они могут; определить только на небольшом расстоянии с помощью боковой линии. Рыбы I могут выделять нужные звуковые сигналы из шумов. Особенно хорошо они различают однотонные звуки, которые подаются ритмично. Центр, регулирующий работу акустико-латеральной системы, расположен в продолговатом мозге, он объединяет работу ядер VII, VIII и X пар нервов.
Электрорецепторная система. Способность животных находить направление своих перемещений во время миграции хорошо известна. Еще в прошлом веке высказывались предположения, что мигрирующие животные могут использовать для целей ориентации и навигации электрические и магнитные поля.
Многие слабоэлектрические рыбы обитают в мутной воде тропических рек, где видимость крайне мала, и они обладают вследствие этого слабо развитым зрением. В условиях ограниченного видения использования для ориентации в пространстве активной электросенсорной системы, включающей в себе электрический орган и систему электрорецепторов. Хорошо развитые у слабо электрических рыб органы слуха, осязания и обычная система боковой линии, также используются в процессе ориентации. Потенциалы достигают 100 мкВ/ см, а их частотных компоненты находятся в диапазоне 0-10 Гц. Функцианальные возможности электрорецепторов перекрывают диапозон электрических полей, создаваемые гидробионтами в естественных условиях.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 806; Нарушение авторских прав?; Мы поможем в написании вашей работы!