КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физиология движения и мускулатура рыб
|
|
|
|
10.1Механизм движения рыб и роль плавников Механическое движение занимает важное место в жизненных процессах. Рыба перемещается в воде — плавает, прокачивает воду сквозь жаберный аппарат, сердце проталкивает кровь по сосудам, кишечник проталкивает пищу. Имеются и другие, менее заметные, но тоже очень важные формы механического движения — изменяется просвет кровеносных сосудов зрачка глаз, сокращаются и растягиваются стенки пузыря, сжимаются и расслабляются кольца
Акт движения рыб осуществляется за счет активной работы плавников. Плавники рыб бывают: парные-грудные, брюшные и непарные-спинной, анальный, хвостовой, спинной плавник может быть один (у карповых), два (у окуневых) и три (у тресковых). Хвостовой плавник принимает некоторое участие в поступательном движении, парализуя тормозящее движение конца тела и ослабляя обратные токи. По характеру действия хвосты рыб принято разделять на: 1) изобатический, где верхняя и нижняя лопасти равновелики: подобный тип хвоста имеется у скумбрии, тунца и многих других: 2) эпибатический – у которого верхняя лопасть развита лучше, чем нижняя: этот хвост облегчает движение вверх подобного рода хвост характерен для акул и осетровых: 3) гипобатический, когда нижняя лопасть хвоста развита больше верхней и способствует движению вниз: такой хвост имеется у летучей рыбы леща и некоторых других рыб. (Рис.66)
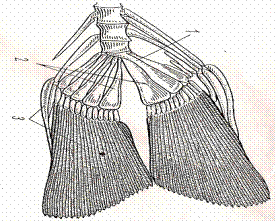
| Рис.65.Хвостовой плавник судака: 1-Конец позвоночного столба-уростиль, 2-гипуралии, 3-кожные и костные лучи-лепидотрихии |
Жировой плавник без костных лучей представляет мягкий кожный вырост на задней части спины (лососевые). Плавники обеспечивают равновесие тела рыб и движения ее в разных направлениях, хвостовой плавник создает движущую силу и исполняет роль руля, обеспечивая маневренность рыбы при поворотах. Спинной и анальный плавники поддерживают нормальное положение тела рыбы, т.е. выполняют роль киля. Парные плавники поддерживают равновесие и являются рулями поворотов и глубин. Основную функцию рулей глубины выполняют у рыб грудные, а также брюшные плавники. С их помощью осуществляется отчасти и поворачивание рыбы в горизонтальной плоскости. Роль непарных плавников сводится к содействию поворотам рыбы вверх и вниз и к роли килей.
На брюшной стороне туловища, каудальное место прикрепления брюшных плавников имеется анальное отверстие, а сразу за ним – мочеполовой сосочек; у части рыб он представляет собой углубление с двумя отдельными отверстиями; мочевым (заднее) и половым. Место расположения заднепроходного, полового и мочевого отверстий за границей тазового и хвостового позвонка.
Плечевой пояс имеет вид хрящевого полукольца, лежащего в мускулатуре стенок тела позади жаберного отдела. Часть пояса называется лопаточным отделом, вентральнее – коракойдным. В основании скелета свободной конечности расположены 3 уплощенные базальные хрящи. Дистальнее базальных хрящей расположены в 3 ряда палочковидные радиальные хрящи. Остальная часть лопасти свободного плавника поддерживается многочисленными тонкими эластиновыми нитями – элементами вторичного кожного скелета.
Тазовый пояс – представлен хрящевой пластинкой, лежащей в толще брюшной мускулатуры перед клоакальной щелью. В брюшных плавниках имеется только один базальный элемент. Он сильно удлинен, и к нему прикрепляется один ряд радиальных хрящей. Остальная часть свободного плавника поддерживается эластиновыми нитями. У самцов удлиненный базальный элемент продолжается за пределы лопасти плавника как скелетная основа копулятивного выроста.
10.1.1 Непарные плавники. Представлены хвостовым анальным и спинным плавниками. У ключей акулы анального плавника нет. Хвостовой плавник акул гетероцеркальный, его верхняя лопасть значительно длиннее нижней, осевой скелет – позвоночник заходит только в верхнюю лопасть.
Скелетную основу хвостового плавника образуют удлиненные верхние и нижние дуги позвонков. Большая часть лопасти хвостового плавника поддерживается эластиновыми нитями. В основании скелета спинных и анальных плавников лежат радиальные хрящи, которые погружены в толщу мускулатуры. Иногда они сливаются в более крупные образования. Свободная лопасть плавника поддерживается эластиновыми нитями. У колючей акулы перед каждым спинным плавником расположен роговой шип, который, как и плакоидные чешуи, представляет собой элемент кожного скелета.
10.1.2 Жаберные крышки. Жаберные дуги представлены каждая четырьмя парными косточками, подвижно сочлененными между собой, и объединяющимися снизу при помощи непарных костных элементов. Пятая, самая задняя жаберная дуга, имеет лишь один парный (нижний) отдел.
Жаберные крышки состоят из четырех вторичных (покровных) костей: крышки (opezculum) и межкрышки (intezopezculum). Жаберная крышка каждой стороны через предкрышечную кость прикрепляется к соответствующей подвеске и к квадратной кости.
10.1.3 Парные конечности и их пояса. Парные конечности представлены грудными и брюшными плавниками. Опорой грудных плавников в теле рыбы служит плечевой пояс. Он представлен двумя небольшими замещающими (первичными) и несколькими покровными костями. Верхняя из замещающих костей – лопатка (scapula) расположена в области причленение свободной конечности. Сразу под ней находится коракоид (coracoideum). Эти два элемента составляют первичный пояс. Они неподвижно соединены с крупной покровной костью клейтрум (coracoideum), верхний конец которой направлен несколько вперед; к нему присоединяется небольшая кость надклейтрум (supracleithum). Клейтрум в свою очередь соединяется с заднетеменной (posttemporale) костью. Направленные вперед нижние концы правого и левого клейтрумов соединяются друг с другом. Позади клейтрума неподалеку от лопатки и коракоида расположена небольшая заднеключичная кость (postcleithrum).
Все названные кости парные, они составляют вторичный плечевой пояс. Правая и левая заднетеменные кости причленяются к осевому черепу, что обеспечивает более прочную фиксацию пояса, и таким ообразом усиливает его опорную функцию.
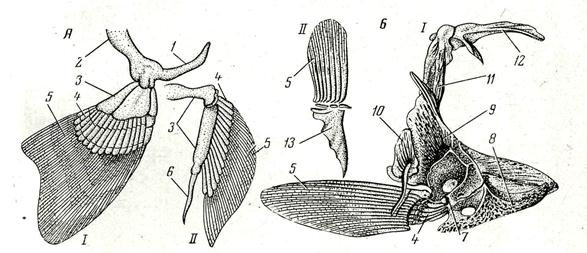
Рис. 66. Скелет парных плавников и их поясов:
А – хрящевая рыба; Б – костистая рыба; 1 – грудной плавник с плечевым поясом;11 – брюшной плавник с тазовым поясом; 1 – лопаточный отдел; 2 – коракоидный отдел; 3 – базалии; 4 – радиалии; 5 – плавниковые лучи; 6 – птеригоподии; 7 –лопатка; 8 – коракоид; 9 – клейт-рум; 10 – задняя клейтрум; 11 – надклейтрум; 12 – задневисочная кость; 13 – тазовая кость
Грудной плавник в своем основании имеет один ряд мелких косточек – радиалий (radialia), отходящих от лопатки (частично и от коракоида). Вся свободная лопасть плавника состоит из членистых кожных лучей (lepidotrichia). Особенность скелета грудных плавников костистых рыб, по сравнению с хрящевыми, заключаются в редукции базалий. Подвижность грудных плавников увеличивается потому, что мышцы прикрепляются к расширенным основаниям кожных лучей, подвижно сочленяющихся с радиалами.
Тазовый пояс представлен сливающимися друг с другом парными плоскими треугольными костями, лежащими в толще брюшной мускулатуры и не связан с осевым скелетом. К боковым сторонам тазового пояса причленяются брюшные плавники. У большинства костистых рыб в скелете брюшных плавников лопасть плавника поддерживается кожными костными лучами (lepidotrichia), расширенные основания, которых непосредственно причленяются к тазовому поясу. Такое упрощение скелета брюшных плавников связано с их ограниченными функциями.
10.1.4 Непарные конечности. Непарные конечности представлены спинными, подхвостовыми (анальным) и хвостовым плавниками. Анальные и спинные плавники состоят из костных лучей, подразделяющихся на внутренние (скрытые в толще мускулатуры) птиригофоры и наружные плавниковые лучи – лепидотрихии.
Хвостовой плавник, имеет влияние равнолопастное строение, однако при рассмотрении его внутреннего скелета видно, что концевые позвонки позвоночного столба сливаются в палочковидную косточку – уростиль (urostal), которая заходит в основание лишь верхней лопасти плавника, а основание нижней лопасти поддерживается расширенными, довольно широкими нижними дугами позвонков гипуралиями. Такой тип строения хвостового плавника, носит название галоцеркального. Наружный скелет хвостового плавника составлен многочисленными кожными лучами – лепидотрихиями.
Из многочисленных способов движения известных среди животных, рыбам свойственны три: плавания, ползания и полет.
В зависимости от выполняемой функции, плавники делят на две группы. Первую группу составляют приспособления, непосредственно направленные на создание локомоторной силы, т.е. морфологические особенности, которые определяются отношением двигателя рыбы. Вторая группа- приспособления, которые движущаяся рыба встречает со стороны определяющей ее виды.
10.2 Функция локомоции и строение хвостового плавника. Функции хвостового плавника многообразны.
1. Хвостовой плавник участвует в общей локомоторной работе туловища, создавая силу, которая толкает рыбу вперед;
2. Действуя, как пассивная несущая полость, итероцеркальный хвостовой плавник создает некоторую подъемную силу, поддерживающую заднюю часть тела рыбы;
3. Хвостовой плавник является стабилизатором и отчасти вертикальным путем.
Хвостовой плавник у большинства рыб способен производить ряд сложных движений. Служит для изменения направления движения в вертикальной плоскости.
Первые три функции могут быть названы пассивными функциями хвостового плавника, так как при отправлении этих функций он действует просто как плоскость, более или менее метко прикрепленная на конце хвостового стебля. Последняя же функция является активной, так как при ее отправлении хвостовой плавник выполняет ряд сложных собственных движений и действует не просто как единая плоскость, а как сложная система, состоящая из отдельных элементов – лучей подвижно скрепленных с концом позвоночного столба. Особенности функции хвостового плавника у различных рыб определяют его форму, в отношении которой должны быть отличены в первую очередь два случая; хвостовой плавник, не разделенный на две лопасти, и хвостовой плавник, раздельный на две лопасти.
Плавание свойственно всем без исключения рыбам. Другие два способа характерны только для немногих видов, причем всегда имеют второстепенное значение. Наиболее общий и типичный для рыб способ движения плавания охватывает собой целый ряд модификации, которые могут быть разделены на две группы:
Первая группа заключает в себе способы плавания, в основе которых лежат волнообразные латеральные изгибания туловища.
Вторая группа – все остальные способы плавания, основу которых составляют те или иные движения плавников по характеру совершаемых плавательных движении делятся на 2 группы. Первая группа плавания основывается на движении плавников неундулирующего типа; преимущественно работа с грудными плавниками, при которых плавник отводится от тела в горизонтальном положении, затем ставится вертикально и приводится к телу, создавая в этот момент определенную локомоторную силу. Способ плавания Chronus chronus (L) u Crenilohrustina (L).Неундулирующие движения плавников являются второстепенным способом движения, при медленном плавании. Основным способом движения оно становится только у некоторых прибрежных медленно плавающих видов. Неундулирующие движения плавников используются в ряде случаев и для передвижения по грунту, как у Gohius иногда и для закапывания в грунт.
Другая группа имеет в своей основе ундулирующие движения плавников спинного и анального. Этот способ плавания встречаются у скатов. Громадному большинству рыб свойственно плавание при помощи латеральных изгибании, этот способ движения является для рыб закономерным. В основе локомоции при помощи ундулирующих движении плавников, лежат волнообразные изгибания пластинки плавника, обусловленные последовательности поперечности отношениями лучей.
Поступательное движение рыб в разных случаях обеспечивается ундуляцией грудных, спинного или анального плавников или же той или иной комбинацией действии этих плавников. У скатов единственным органом локомоции являются чрезвычайно расширенные грудные плавники. Большая поверхность этих плавников и значительная дорзовентральная подвижность их лучей превращает их в прекрасный ундулирующий аппарат. Волны ундуляции приходят по плавникам спереди назад, создавая движущую силу, направленную вперед. Дублирующими мембранами, сообщающими рыбе поступательные движения, являются спинной и анальные плавники. В этом случае совокупное действие этих плавников создает силу, движущую рыбу вперед, причем волны ундуляции на плавниках направлены параллельно линиям основания плавника. Благодаря волнообразным изгибаниям пластинки плавника, возникают силы, толкающие рыбу вперед.
10.2.1 Функции плавников. В теле рыбы выделяются четыре постоянные зоны, для которых специфичны определенные функции плавников. Эти зоны можно назвать функционально специфическими, так как для каждой из них специфична определенная функция или ряд функций. I - зона передних рулей, и несущих плоскостей, II - зона килей, III-зона стабилизаторов, и IV- зона задних рулей и локомоторных органов. Функции зоны I выполняются парными плавниками брюшных плавников или только грудными.
Функции зоны II выполняются той частью спинного плавника, которая расположена спереди от центра тяжести, а также брюшными плавниками и отчасти анальным, если эти плавники или их передние отделы расположены спереди от центра тяжести. Функции зоны II выполняются морфологическими обособленными отделами спинного и анального плавников, которые расположены сразу за центром тяжести.
Функции зоны III несут самые задние отделы спинного и анального плавников (если они приближены к заднему концу тела) и хвостовой плавник.
Указанное расположение на теле функционально специфических зон характерны для всех рыб вписывающих при помощи волнообразных изгибании тела.
10.3 Плавание рыб. Плавание рыб происходит благодаря работе поперечнополосатой мускулатуры, которая приводит в движение движительные плавники. Рыбы передвигаются в толще воды, создавая упор, т. е. отталкиваясь от среды, обладающей некоторой плотностью, вязкостью и податливостью. Проще всего понять явление упора, рассматривая гребное плавание при помощи парных грудных плавников.
Грудной плавник, закрепленный своим основанием на плечевом поясе, под действием мускулатуры совершает движения, подобные вращению калитки вокруг петель. При движении назад, во время гребка, плавник максимально расправлен и создает максимальный упор. При движении плавника вперед, при подготовке гребка, лучи сведены и создают минимальный тормозящий упор. Гребным плаванием пользуются щуки, зеленухи, колюшки, бычки, сростночелюстные и многие другие рыбы.
Одиночный цикл движения плавника может быть подразделен на движение вперед (подготовка гребка), движение назад (гребок) и период, когда плавник прижат к телу (рефрактерная фаза). Скорость плавания рыбы зависит от скорости движения плавника — скорость движения внешнего края плавника всегда больше скорости поступательного движения рыбы, т. е. плавник является двигателем. Угловая амплитуда движения плавника может достигать 900. Скорость растет при уменьшении доли времени рефрактерной фазы и при росте доли времени активных фаз — подготовки и гребка, которые занимают приблизительно одинаковую часть времени. Скорость плавания находится в прямой зависимости от частоты гребков, которая может достигать 20 Гц, но обычно не превышает 5 Гц.
Движение грудных плавников у большинства рыб поочередное (типа «кроль»), но у некоторых видов, например у карпа,— одновременное (типа «брасс»). Скорость плавания при работе одних только плавников небольшая, расход энергии тоже, по-видимому, небольшой, так как в работу вовлекаются небольшие массы мускулатуры плечевого пояса. Коэффициент полезного действия мускулатуры при таком типе плавания довольно высок.
При плавниковом плавании туловищно-хвостовая мускулатура не бездействует, она в какой-то степени напряжена для поддержания обтекаемой «позы». В противном случае тело начинает пассивно колебаться в набегающем потоке, как водоросль в потоке, или как флаг на ветру (так называемый флаттер), и это вызывает торможение.
Ограниченные массы мускулатуры вовлекаются в работу при плавании с помощью непарных плавников, например спинного, анального, когда эффективный упор создают поперечные складки плавников. Наибольшие скорости достигаются рыбами при периодическом волнообразном изгибании всего тела (ундуляции). Движителем в этом случае служит почти вся поверхность тела, за исключением негибкой головы.
При движении рыб активно сокращается почти вся туловищно-хвостовая мускулатура, составляющая приблизительно половину массы тела. Упор при ундуляционном плавании создается искривлением тела и движением локомоторной волны от головы к хвосту. За один цикл движения рыба могла бы продвинуться на длину тела до теоретического финиша, но в результате наличия КПД движителя 0,70 возникает пробуксовка и она проплывает расстояние до фактического финиша.
Н. В. Кокшайский охарактеризовал плавание как явление, при котором организм образует некоторую систему подвижных элементов, отталкивающих среду и одновременно отталкивающихся от нее. Длина локомоторной волны (расстояние между соседними гребнями) соизмерима с длиной рыбы.

Рис.67. Схематическое изображение плавания рыбы. Стрелки изображают направление приложения упора.
Скорость движения локомоторной волны по телу рыбы равна произведению длины волны на частоту взмахов хвоста, численно она также близка произведению длины рыбы на частоту ундуляции:
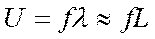
где U — скорость локомоторной полны; f—частота ундуляции; 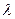 — длина локомоторной волны; L — длина рыбы. Вода представляет собой податливую среду, поэтому в ней имеет место некоторая «пробуксовка» движителей. Скорость плавания рыбы поэтому всегда меньше скорости локомоторной волны:
— длина локомоторной волны; L — длина рыбы. Вода представляет собой податливую среду, поэтому в ней имеет место некоторая «пробуксовка» движителей. Скорость плавания рыбы поэтому всегда меньше скорости локомоторной волны:
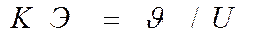
где КЭ — коэффициент эффективности движителя; 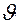 — скорость плавания рыбы.
— скорость плавания рыбы.
Максимальная скорость плавания рыбы является функцией длины тела и максимальной частоты ундуляции, т. е. максимальной частоты сокращений туловищной мускулатуры. Максимальная частота ударов хвоста при плавании некоторых достигает 30 Гц.(Таблица 20).
Максимальные частоты ундуляции и соответствующие им максимальные, так называемые спринтерские скорости могут поддерживаться в течение нескольких секунд. Далее наступает утомление и скорость резко падает, это связано со строением рыбы: с большим числом позвонков могут сильнее изгибать тело, чем рыбы с малым числом позвонков. Число позвонков у рыб колеблется поразному: так у рыбы -луны от 16 и у ремень рыбы до 400. Рыбы с мелкой чешуей также могут изгибать свое тело в большей степени, чем крупночешуйные.
Таблица 21 - Расчетные значения скорости плавания рыб (см./с) как функция частоты ундуляции (Гц) и длины рыбы (см) для области чисел Рейнольдса менее 105 (по Яржомбек, 1975)
| Длина рыбы | Частота ундуляции | |||||||||
| 0,1 1,2 5,7 | 0,7 3,4 | 0,2 2,4 8,4 | 0,7 4,5 | 1,3 6,8 | 2,1 9,2 | 4,8 | 6,9 | 9,6 |
Скорость движения рыб связана и с биологическим состоянием рыбы, в частности зрелостью гонад, температурой воды может меняться и от того, движется рыба в стае или в одиночку. Наибольших скоростей достигают некоторые акулы, меч- рыбы, тунцы. Голубая акула перемещается со скоростью около - 10 м./сек, тунец со скоростью 20м/ сек, лосось 5м/сек. Абсолютная скорость движения рыбы зависит от ее размеров. Крупные рыбы обладают большей скоростью, чем более мелкие. Эта зависимость выражается следующей формулой:
V = 1 [ L (3f-4)]
Где V - скорость в см./ сек.
L - длина рыбы в см.
f - число взмахов хвоста в секунду
Поэтому для сравнения скорости движения разноразмерных рыб используется обычно коэффициент скорости представляющий собой частное деление абсолютной скорости движения рыбы на корень квадратный ее длины (V).
Рыбы хорошие пловцы, в текучей воде по форме тела отличаются от хороших пловцов в стоячей воде, в частности у первых хвостовой стебель обычно значительно выше и короче, чем у вторых. В качестве примера можно сравнить форму хвостового стебля форели, приспособленной к жизни в воде с быстрым течением, и скумбрии- обитателя медленно двигающихся и стоячих морских вод.
Быстро плавая, преодолевая быстрины и перекаты, рыбы утомляются. Они не могут длительно плавать без отдыха. При большом напряжении у рыб в крови накапливается молочная кислота, которая после отдыха исчезает. Молочная кислота влияет на растворимость газов кровью и на транспортную функцию гемоглобина. Рыбы, при прохождении рыбоходов, утомляются, преодолевая их, даже гибнут.
Благодаря специфическому строению мускулатур рыб, сокращение каждого миомера вызывает изгиб тела на довольно большой протяженности, т. е. создается рычаг приложения силы. Мышечные волокна в миомерах ориентированы таким образом, что одно волокно является как бы продолжением другого и такие составные нити косо расходятся от средней линии по направлению к хвосту и краям тела.
Светлая и темная мускулатура рыб во многом различна. При спокойном плавании рыбы в крейсерском режиме число сокращений волокон темной мускулатуры равно числу плавательных движений. Сокращения волокон белой мускулатуры носят нерегулярный характер.
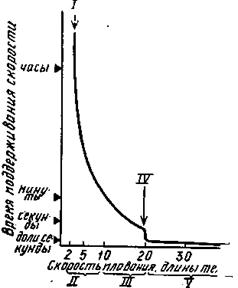 Существует некоторая неутомляющая частота мышечных сокращений, при которой времени между двигательными актами достаточно для полного восстановления работоспособности. При таких режимах движения происходят длительные миграции рыб. Неутомляющая средняя скорость в 5—10 раз меньше максимальной спринтерской скорости плавания. Проведенные эксперименты и наблюдения в естественных условиях показали, что лососи не утомляются при скорости плавания 1—2 длины тела в секунду, т. е. до 5 км/ч.
Существует некоторая неутомляющая частота мышечных сокращений, при которой времени между двигательными актами достаточно для полного восстановления работоспособности. При таких режимах движения происходят длительные миграции рыб. Неутомляющая средняя скорость в 5—10 раз меньше максимальной спринтерской скорости плавания. Проведенные эксперименты и наблюдения в естественных условиях показали, что лососи не утомляются при скорости плавания 1—2 длины тела в секунду, т. е. до 5 км/ч.
Рис.68. Скорости плавания быстроходных рыб: I- неутомляющая скорость;II- стайерская скорость; III- скорость средних дистанции; IV- спринтерская скорость; V- скорость прыжков с разбега
Тунцы — рекордсмены среди рыб; по скоростной выносливости могут длительное время двигаться со скоростью 3—4 длины тела в секунду, т. е. порядка 20 км/ч.
Между неутомляющими и спринтерскими скоростями имеется область, где время поддерживания скорости находится в обратной связи с заданной скоростью.
По аналогии со спортивной терминологией эту область можно разбить на «средние дистанции» — до времени движения несколько десятков минут и «стайерские дистанции». Стайерская скорость может поддерживаться несколько часов, но она все-таки не является неутомляющей, или крейсерской, скоростью (рис. 69).
Наилучшие среди рыб пловцы — тунцы, ставриды, лососи. Они плавают в 2-3 раза быстрее и выносливее, чем посредственные пловцы, такие, как осётровые, камбалы, бычки, угри.
Таблица 22 - Максимальная частота плавательных движений
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 6512; Нарушение авторских прав?; Мы поможем в написании вашей работы!