КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Долговременная потенциация
|
|
|
|
ЦАМФ
(циклический нуклеотид)
_А-
| Протеинкиназа А |
| Протеинкиназа С |
| Фосфорилирование белков Са^-каналов |
| Фосфорилирование белков К-каналов |
| Открытие потенциал-независимых Са2"1" -каналов |
| Закрытие К-каналов |
Рис. 30. Основные этапы фосфорилирования Са^- и К-каналов, обеспечивающего кратковременное изменение проводимости синапса.
ких процессов синтезируется циклический аденозинмонофосфат (цАМФ). За счет двойного действия на аденилатциклазу Са2^ и серотонина возрастает образование цАМФ. Активированные киназы фосфорилируют белки ионных каналов. Киназа А взаимодействует с белками кальциевых каналов и открывает их для входа ионов кальция. Киназа С фосфорилирует белки калиевых каналов, вызывая
| Повторное раздражение головы |
| Протеинкиназа фосфорилирует калиевые каналы и ипактивируст их |
|
| Регистрация калиевого тока через изолированный кусочек мембраны |
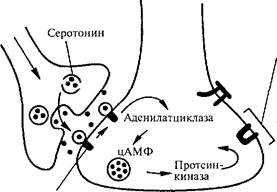
Серотонин активирует рецепторы
Рис. 31. Зависимое от активности пресинаптическое облегчение у аплизии (по Э. Кэнделу и Р. Хокинсу, 1992).
их закрытие. Это уменьшает калиевый ток из нейрона и калиевую гиперполяризацию, следующую за ПД и ВПСП, что приводит к удлинению ПД. На рис. 31 показана цепочка реакций, приводящих к инактивации К-каналов, вызываемой возбуждением модулирующих нейронов. Продление ПД удерживает потенциал-зависимые Са^-каналы в открытом состоянии, что увеличивает в нейроне содержание свободного кальция и выброс медиатора. Фосфорили-рование калиевых каналов (8-каналов) рассматривается как одна из форм кратковременной памяти, которая ограничена временем жизни белка, образующего 5-каналы, и легко стирается при воздействии фосфопротеинфосфотаз (дефосфорилирующих 5-каналы).
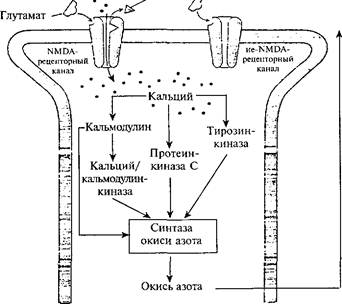
События, разворачивающиеся на постсинаптическом нейроне у млекопитающих в ситуации обучения, связаны с особой группой рецепторов, медиатором для которой служит глутамат. Среди них различают NМ^А-рецепторы (они селективно активируются синтетическим аналогом глутамата Т^-метил-0-аспартатом) и не- NМ^А-рецепторы. Каналы, связанные с NМ^А-рецепторами, имеют двойные ворота, открывающиеся при выполнении двух условий: мембрана должна быть сильно деполяризована, а к рецептору должен присоединиться медиатор глутамат (рис. 32). Каналы, связанные с ММОА-рецепторами, находясь в открытом состоянии, пропускают очень большой поток ионов Са2"*". Эта их отличительная особенность очень важна для механизма памяти. Не-КМОА-рецепторы, реагируя на глутамат, сразу открывают кальциевые
| Пресинапти-чсскии нейрон Кальций Пузырек 'уГ\ с нейромедиа- тором |
|
| Усиленное выделение нейромедиатора АОР-Рибозил- |
| Участок выделения нейромедиатора |
| трансфераза сОМР <—Гуанилат-< * циклаза |
| АОАОАОАОАОАОА |
| Кальций •. Магний /. ^ ^ |
|
| Постеннапти-ческий нейрон |
|
| Ретроградный мессенджер (окись азота) |
Рис. 32. Внутриклеточные процессы в постсинаптическом нейроне гиппо-кампа, реагирующего на глутамат (по Э. Кэнделу, Р. Хокинсу, 1992).
ионные каналы без дополнительных условий и Са^ входит в нейрон. Это создает уровень деполяризации нейрона, который необходим для реакции г^МОА-рецепторов. Деполяризация нейрона снимает блокаду с гШОА-каналов, которая осуществляется внеклеточным магнием (Мё). Изменения на посте инаптической мембране с участием NМ^А- и не^МОА-рецепторов наиболее хорошо изучены у нейронов гиппокампа на модели долговременной
10-3341
Привыкание
| Многократное раздражение сифона |
| Раздражение сифона |
| Потенциал-зависимые Са^-каналы и н а кти в иру юге я |
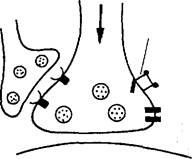
|
| Количество выделяемого медиатора снижается |
Слабая реакция мотонейрона
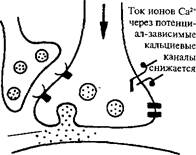
| Сенситизация Повторное раздражениеголовы |
| Раздражение сифона |
|
| Калиевые» каналы инактиви-руются |
| Количество выдел яемого /^''.'. ^медиатора возрастает |
| Продолжительное поступление Са2* вследствие того, что инактивация калиевыхканалов увеличивает длительностьпотенциала действия |
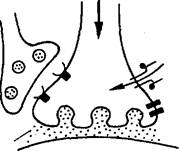
Сильная реакция мотонейрона
Рис. 33. Механизм передачи сигнала от сенсорного нейрона к моторному, участвующего во втягивании жабры у аплизии при привыкании (а) и син-ситизации (б) (по Б. Альбертсу и др., 1994).
потенциации. Показано, что NМ^А-рецепторы играют ключевую роль в долговременной потенциации. Если такие каналы селективно блокировать с помощью специального ингибитора, то долговременная потенциация не происходит, хотя синаптическая передача не нарушается. При этом животное теряет способность к тому виду научения, который связан с гиппокампом. Искусственно регули-146
руя внутриклеточный уровень ионов Са24', можно усилить потен-циацию или предотвратить ее. Ионы Са24', войдя в постсинапти" ческуто клетку, соединяются с кальмодулином и в конечном счете активируют зависимые от него киназы. Предполагается, что кина-зы стимулируют синтез ретроградного мессенджера, который проникает в пресинаптический нейрон и там дополнительно усиливает выброс из него медиатора глутамата.
Ионные процессы при сенситизации и привыкании в системе сенсорный нейрон — моторный нейрон, отвечающей за исполнение рефлекса втягивания жабры у аплизии (по результатам опытов Э. Кэндела), схематически представлены на рис. 33. При сенситиза-дии, которую можно вызвать у моллюска ударами по голове, активируются модулирующие нейроны. Они образуют на пресинап-тических окончаниях сенсорного нейрона синапсы, выделяющие серотонин. Серотонин через аденилатциклазу, с которой он взаимодействует, приводит к инактивации калиевых каналов, что создает условия для удлинения ПД у сенсорного нейрона. В результате сенсорный нейрон может выделять больше нейромедиатора в синаптическую щель, чем обычно. Аналогичный эффект сенсити-зации у этих же нейронов можно получить, прикладывая серотонин непосредственно к их мембране.
При привыкании повторяющееся возбуждение сенсорных клеток (на раздражение сифона) изменяет белки ионных каналов таким образом, что приток Са24" в клетку падает за счет инактивации кальциевых каналов и количество высвобождаемого медиатора в синаптическую щель уменьшается.
Рассмотренные выше привыкание и сенситизация — очень простые формы научения. При ассоциативном научении процессы, происходящие в нервных клетках, значительно более сложны. Однако нейронные механизмы пластичности и связанные с ними ионные процессы, определяющие сенситизацию и привыкание, используются и в системе ассоциативного обучения. Так, события, развертывающиеся в нейроне во время сенситизации, могут быть обнаружены на начальном этапе ассоциативного обучения нейрона.
Отличительной особенностью ассоциативного обучения является наличие подкрепления, в роли которого может выступать сильное безусловное раздражение- Формирование ассоциации у нейронов предполагает наличие конвергенции подкрепляющего раздражителя и будущего условного сигнала. Механизмы условного рефлекса детально были изучены на нейронах моллюсков. Особенно следует отметить вклад, который внесли работы Э. Кэндела и его коллег, выполненные на морском моллюске аплизии. У моллюска имеется простой безусловный оборонительный рефлекс в
10-
виде втягивания жабры, служащей органом дыхания, в ответ на раздражение его тела с противоположной стороны от жабры (сифона — мясистого отростка). Рефлекс вызывается возбуждением сенсорных нейронов сифона, которые прямо контактируют с мотонейронами жабры, а также с рядом интернейронов, образующих синапсы на тех же мотонейронах. Кроме того, рефлекс может быть вызван раздражением и других частей тела, в частности «хвоста», электрическая стимуляция которого и была использована в качестве безусловного раздражителя при выработке условного рефлекса. Установив, что у моллюска можно выработать условный оборонительный рефлекс — втягивание жабры — на слабое тактильное раздражение сифона, подкрепляемое электрическим ударом по «хвосту», авторы исследовали связи между сенсорными нейронами сифона и их клетками-мишенями — мотонейронами и интернейронами. Было обнаружено, что после выработки условного рефлекса слабая стимуляция сенсорных нейронов, связанных с сифоном, одновременно начинает вызывать возбудительные синап-тические потенциалы как в интернейронах, так и в моторных, что и определяет появление условнорефлекторного втягивания жабры.
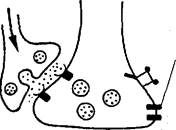
Э, Кэндел и Р. Хокинс (1992) так описывают результаты своих исследований по выработке условного оборонительного рефлекса у аплизии. Безусловный раздражитель — электрический удар по «хвосту» — активирует модулирующие нейроны, выделяющие се-ротонин. Таким образом, безусловный раздражитель представлен в сенсорном нейроне опосредованно через модулирующие нейроны-Условный раздражитель на сенсорном нейроне представлен их собственной активностью. Пунктом их конвергенции в сенсорном нейроне является аденилатциклаза, активация которой через два входа создает эффект, аналогичный сенситизации. Повышение проводимости через синапс в пресинаптическом нейроне, возникающее при ассоциативном обучении за счет совпадения активирующих влияний, поступающих через разные входы, требует от них соблюдения определенных временных отношений. Условный сигнал должен опережать действие безусловного на некоторый критический интервал времени. Это правило временных отношений условного и безусловного раздражителей было сформулировано И.П. Павловым. Для условного рефлекса втягивания жабры время опережения условным сигналом безусловного составляет около 0,5 с. Модулирующие нейроны, активированные болевым раздражением «хвоста» и действующие как усилители, вызывают большее пре-синаптическое облегчение сенсорных нейронов, если последние незадолго до этого находились в активном состоянии. Стимуляция же одних модулирующих нейронов вызывает меньшее синапти-
| Пресинапти-ческий нейрон |
| Пресинапти-ческий нейрон |
|
| Модулирующийнейрон |
|
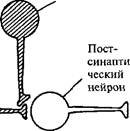
| Пост- | |
| сил апти- | Пресинапти- |
| ческий | ческий нейрон |
| / нейрон | |
| ^|. 1 | |
| ^^"Ц |
|
Рис. 34. Два принципа обучения: пре-постсинаптическое совпадение — синапс Хебба (а) и обучение, зависимое от модулирующего нейрона, — пре-модулирующее совпадение, или зависимое от активности облегчение (б) (поЭ. Кэнделу, Р. Хокинсу, 1992).
ческое облегчение. Поэтому необходимо опережение безусловного раздражителя активацией пресинаптического нейрона, которая представляет действие условного раздражителя.
Это свойство пресинаптического облегчения, зависящее от вклада модулирующих нейронов, в 1963 г. Л. Тауц и Э. Кэндел назвали обучением, зависящим от активности (рис. 34), и стали его рассматривать как новый принцип обучения. Поэтому к первому принципу обучения, предложенному в 1949 г. Д. Хеббом, добавился второй. Принцип Д. Хебба утверждает необходимость совпадения активности пресинаптического и постсинаптического нейронов как условие усиления связи между ними (пре-постсинаптическое совпадение). Он получил название «синапс Хебба^. Согласно второму принципу для усиления связи между нейронами активность постсинаптической клетки не нужна, если одновременно с пресинаптической клеткой активирован третий нейрон — модулирующий (пре-модулирующее совпадение).
Суть обучения, зависящего от активности, составляет одновременная активация фермента аденилатциклазы ионами Са2^, входящими в нейрон во время генерации ПД, и серотонином от модулирующих нейронов.
Появление пресинаптического облегчения в составе ассоциативного обучения за счет участия модулирующих нейронов доказано при формировании условного оборонительного рефлекса у виноградной улитки. В системе оборонительного рефлекса улитки модулирующие нейроны представлены пулом серотонинергичес-
ких клеток педального ганглия. Их модулирующее влияние широко распространяется в том числе и на пресипаптические окончания сенсорных нейронов на командном нейроне оборонительного рефлекса. Избирательная блокада модулирующих нейронов препятствует выработке условного рефлекса. Сопровождение условного сигнала ударом тока активирует модулирующие нейроны, которые запускают механизм пресинаптического облегчения.
Неясно, как далеко можно экстраполировать данные, полученные на моллюсках. Таков ли клеточный механизм фиксации следов памяти и у других животных? Э. Кэндел и Р. Хокинс полагают, что при обучении у моллюска и у млекопитающих действуют сходные механизмы, использующие модулирующее вещество, вызывающее зависимое от активности усиленное выделение ней-ромедиатора из пресинаптического нейрона. По их мнению, долговременная потенциация в гиппокампе как одна из форм обучения млекопитающих требует участия специального гипотетического модулирующего вещества,
|
|
|
|
|
Дата добавления: 2014-12-10; Просмотров: 1277; Нарушение авторских прав?; Мы поможем в написании вашей работы!