КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Молекулярные механизмы пластичности
|
|
|
|
Повышенный интерес к изучению молекулярных механизмов памяти в 50—60-х годах был вызван работами известного шведского нейрохимика X. Хидена, изучавшего роль рибонуклеиновой кислоты (РНК) в процессах памяти и положившего начало исследованию кодирования приобретаемого навыка последовательностью нуклеотидов РНК. Он изучал содержание РНК и белков в нейронах вестибулярной системы у обученных крыс и кроликов. Крыс тренировали взбираться по наклонно натянутой проволоке, к концу которой прикреплялась приманка- Кролики вращались в устройстве, напоминавшим детскую карусель. Оказалось, что освоение животными нового поведения сопровождалось ростом синтеза РНК и белков. Предполагалось, что последующий синтез белка на матрице «обученной» РНК сохраняет приобретенную информацию («запоминающие белки»). Гипотеза X. Хидена о роли РНК в кодировании памяти была критически переоценена самим автором и не получила подтверждения опытами других исследователей. В серии работ, выполненных многими авторами,
была установлена связь обучения с ростом синтеза ядерной и ци-топлазматической РНК во многих структурах мозга. Его блокада с помощью рибонуклеазы, вводимой разными способами (в структуры мозга, на его поверхность или внутрибрюшинно), у крыс и кроликов приводила к исчезновению ранее выработанных условных рефлексов и затрудняла обучение новым навыкам. Однако до сих пор не доказана биохимическая специфичность белков, синтезируемых в процессе запоминания. Поэтому роль РНК и белков в механизмах памяти исчерпывается их участием в восстановительных процессах клеток, вовлеченных в активное функционирование. Позднее, в 70-х годах, исследователи пришли к выводу о том, что сохранение приобретенной информации связано не с РНК, а с ДНК.
Существенный вклад в понимание молекулярных механизмов памяти внесли Франсуа Жакоб и Жак Моно, которые во Франции в 1961 г. впервые опубликовали результаты своих исследований белкового синтеза у бактерий. Они описали сложную структуру генетического контроля со специальным механизмом, обеспечивающим адаптацию организма к воздействиям внешней среды. За эту работу, признанную теперь классической, они были удостоены впоследствии Нобелевской премии. Обычно бактерия использует в качестве источника углерода фруктозу или сахарозу. Но если ее перенести на среду, содержащую сахар лактозу, которую нужно предварительно расщепить, чтобы получить углерод, то часть молекул лактозы, проникая в клетку, запускает специальный механизм, который изменяет репрессор — белок, подавляющий транскрипцию ДНК в РНК. Это делает возможным синтез фермента, необходимого для расщепления лактозы. Действие реп-рессора можно устранить с помощью других белков или гормонов. Таким образом, помимо структурных генов, хранящих информацию об организме, они выделили особые регуляторные белки, которые, присоединяясь к ДНК или отделяясь от нее, контролируют экспрессию генов.
Существует много моделей, объясняющих связь памяти с генетическими структурами нейрона. Большинство из них утверждает наличие единого процесса экспрессии генов, стимулируемого обучением. Она ведет к синтезу многих классов белков — рибосом-ных, синаптических структурных белков аксонных окончаний, энзимов (ферментов) для синтеза мембранных липидов.
Двухфазная модель синтеза белков как основа долговременной памяти впервые была предложена Э. Глассманом (СИавхтап Е., 1969). Опираясь на результаты опытов других исследователей, показавших, что ингибитор синтеза белка у золотой рыбки одно-
временно вызывает блокаду синтеза РНК и дефицит памяти, он описал цепь молекулярных событий, определяющих долговременное хранение информации, как последовательность: белок-1 ~ РНК — белок-2. Э. Глассман предположил, что белок-1, синтезируемый во время обучения, является активатором для специфических генов, которые на следующем шаге кодируют РНК, опре-делящую синтез белка-2. Последний причастен к консолидации памяти. В результате синаптические связи, которые появляются между нейронами в процессе обучения, превращаются в постоянные.
Идеи Э. Глассмана получили подтверждение в серии работ X. Мэттиаса и его коллег (МаНЫев Н. еТ: а1., 1989), обративших внимание на наличие двух волн активации синтеза белков после обучения крыс различению стимулов. Первая волна белкового синтеза начиналась непосредственно после тренировки, в то время как вторая появлялась 6—8 ч спустя. Был сделан вывод, что две стадии формирования долговременной памяти требуют разных белков: регуляторных и эффекторных соответственно.
Экспериментальные доказательства двух фаз активации синтеза РНК и белков при обучении крыс получены с помощью радиоактивных методов. Регулярное измерение включенных в РНК и белки меченых предшественников в разные сроки после обучения обнаружило отставленные во времени два пика активации транскрипции и трансляции. При обучении крыс активному избеганию содержание радиоактивных РНК и белка возрастает в течение первого часа после сеанса обучения и вторично на 6—10-м часу.
А. Барзилаи (ВаггПа! А. е1 а1., 1989) с коллегами из лаборатории Э, Кэндела описали две волны активации синтеза белка у сенсорных нейронов морского моллюска аплизии, когда долговременное синаптическое облегчение создавалось (в течение более 24 ч) аппликацией серотонина в течение 1,5 ч или 2-часовой аппликацией аналога цАМФ. Активация синтеза белка измерялась по включению радиоактивного "З-метионина. Появление первой волны активации синтеза белка наблюдалось через час после подведения серотонина, ко 2-му часу она исчезала. На 3-м часу возникала вторая волна активации синтеза белка, более выраженная и протяженная во времени.
Анализ состава синтезируемых белков с помощью гель-электрофореза показал, что на первой фазе активации под влиянием серотонина и цАМФ создаются одни и те же 15 белков. Их активация кратковременна (от 1 до 3 ч) и наступает через 15—30 мин после начала воздействия. Вторая волна появляется через 3 ч после начала воздействия и длится около 5 ч; она включает синтез четырех новых белков, а через 24 ч — синтез еще двух.
Одна из популярных сегодня молекулярных моделей памяти представляет процесс формирования памяти следующим образом. Внешнее воздействие (изменение экстраклеточной среды) вызывает в геноме нейрона каскадную реакцию, в которой выделяются две фазы активации синтеза белков и РНК. Первая фаза активации соответствует индукции специфических регуляторных генов из класса «непосредственно ранних генов». Продукты ранних генов индуцируют экспрессию «поздних» генов — морфорегуляторных генов. Они определяют вторую фазу активации синтеза РНК и белков, что ведет к росту и/или изменению клеточных связей в мозге.
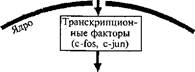
«Ранние гены» впервые были обнаружены при изучении влияния фактора роста на нейроны в культуре ткани. По своим свойствам они напоминали «непосредственно ранние гены» бактериофагов и эукариотических ДНК вирусов. Поэтому по аналогии с вирусными генами эти быстро активирующиеся клеточные гены были названы «генами первичного ответа», «генами раннего ответа», «генами компетентности». Одним из первых ранних генов, который был идентифицирован и клонирован, стал ген с-Го5. Предполагается, что он контролирует клеточный рост и пролиферацию. К настоящему времени клонировано уже около 100 ранних генов. Продукты большинства ранних генов представляют собой регуля-торные белки. Ранние гены контролируют транскрипцию поздних генов, являющихся для них мишенями. Они влияют на регулятор-ные элементы поздних генов в их промоторных областях, которые служат рецепторами для продуктов ранних генов.
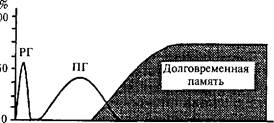
Механизм долговременной памяти с учетом двухкаскадной реакции экспрессии генов представлен на рис. 36. Показаны волны экспрессии ранних генов (РГ) и поздних генов (ПГ). ДП формируется после экспрессии поздних генов.
В ряде работ специально проверялась связь первой волны активации синтеза РНК и белков с экспрессией ранних генов. Индукция ранних генов (с-Гоз, с-]ип), контролируемая с помощью введения радиоактивных предшественников РНК, была обнаружена в неокортексе мышей через 15 мин уже после одного опыта с обучением пассивному избеганию. Сходный эффект был зафиксирован через 30 мин после сеанса обучения крыс активному избеганию электрического тока (запрыгиванием на полочку): синтез мРНК возрастал вследствие индукции раннего гена с-Го5, но не с-тус. Изменение наблюдали в коре, гиппокампе и мозжечке (Анохин К.В„ 1997).
Экспрессия ранних генов возникает на ранней стадии обучения и с автоматизацией навыка исчезает (Анохин К.В., 1997). Чем труднее идет обучение, тем сильнее выражена их экспрессия. Хуже
0246 12 Время (часы)
| Обучение |
| Ранние гены |

|
Рис. 36. Две волны экспрессии генов, определяющих появление долговременной памяти (ДП).
Экспрессии ранних (РГ) и поздних (ПГ) генов. На абсциссе — время после обучения; на ординате — эффективность процессов, определяющих долговременную память (по К.В. Анохину, 1997).
обучающиеся мыши (неуспешная группа) отличались в 1,8 раза большей экспрессией гена с-Гоа по сравнению с успешной группой. Экспрессия ранних генов с-Гок и с-)ип стимулировалась не только ситуацией обучения, но и помещением животного в новую, сенсорно-обогащенную среду, а также отменой ожидаемого отрицательного подкрепления.
Удобным объектом для изучения механизмов памяти всегда был гиппокамп. На нем легко можно смоделировать процесс обучения, создавая в нем электрической стимуляцией эпилептический очаг активности. С помощью измерения синтеза РНК радиологическими методами было установлено, что эпилептическая активность увеличивает экспрессию ранних генов в пирамидных клетках гип- покампа и зубчатой фасции. Кроме того, в этих же структурах гип-покампа наблюдается экспрессия двух других генов. Один кодирует синтез протеинкиназы А, ответственной за фосфорилирование белков. Экспрессия другого гена вызывает синтез рецепторов ММОА, медиатором для которых служит глутамат. Известно, что через эти рецепторы в клетку входит особенно много ионов Са24", поэтому они непосредственно участвуют в процессе обучения.
Сильное влияние на представление о молекулярных механизмах памяти оказали работы Э. Кэндела и его коллег (Сое1е1 Р. е1 а1., 1986; Кагк1е1 Е. е1 а1., 1987). Его теория каскада молекулярных реакций при обучении предполагает три уровня памяти, Кратковремен-
пая память, которая длится от нескольких минут до нескольких часов, обусловлена ковалентной модификацией белков, не достигших состояния возбуждения. Промежуточная память, охватывающая несколько часов, обусловлена фосфорилированием белков. Долговременная память, длящаяся более чем один день, зависит уже от индукции новых генов через вторичных посредников и регуляторов транскрипции. Авторы предположили, что ранние эф-фекторные гены ответственны за синтез белков, которые сохраняют память в течение дней. Память, сохраняемая в течение недель и месяцев, поддерживается другими — поздними эффекторными генами, которые включаются ранними регуляторными генами.
На основе накопленных за последние годы сведений об экспрессии генов при обучении некоторые исследователи приходят к выводу о том, что память использует тот же генетический аппарат, который связан с онтогенетическим развитием организма, с регуляцией роста и дифференциацией его клеток. Подчеркивается общность молекулярного каскада экспрессии генов при обучении и развитии. Оба процесса реагируют на изменения экстраклеточной среды. На стадии развития организма новые экстраклеточные сигналы ведут к инициации экспрессии генов, обеспечивающей клеточную дифференцировку и консолидацию генетически запрограммированных функциональных систем.
При обучении комбинация экстраклеточных сигналов вызывает реэкспрессию генов, бывших активными на стадии созревания и онтогенетического развития организма.
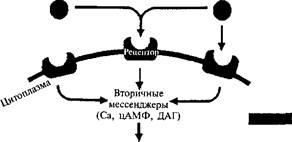
На рис. 37 показано сходство основных этапов процессов экспрессии генов при развитии и обучении. Экстраклеточные сигналы активируют вторичные мессенджеры, которые высвобождают каталитические субъединицы протеинкиназ. Протеинкиназа транспортируется в ядро клетки и там через транскрипционные факторы действует на ранние гены (с-Гох и с-]ип), вызывая их экспрессию, Последние кодируют транскрипционные факторы для поздних генов — белки: РОЗ и ЗШ. Эти белки и вызывают экспрессию поздних генов, которая определяет синтез различных структурных белков и молекул, необходимых для синаптического роста.
Рассмотренные клеточные и молекулярные механизмы обучения не касаются проблемы специфичности этого процесса. Однако при обучении синаптическое облегчение проведения сигнала возникает селективно, т.е. на отдельных синапсах. Для объяснения этого явления Т.Н. Греченко и Е.Н. Соколов (1987) предположили, что вся мембрана нейрона проецируется на геном. Продукты реакций отдельных локусов мембраны, вызванные экстраклеточным сигналом в виде белка-регулятора (БР), стимулируют экспрессию или
РАЗВИТИЕ ОБУЧЕНИЕ
Внеклеточные сигналы
| Трофические факторы |
| Протеин киназы (РКС,РКА) |
Экспрессия кгенов-мишеней»" (1Ч-САМ, ОАР-43)
| Консолидация^ функциональных систем / |
| Модификация^ функциональных систем ^ |
Рис. 37. Общность молекулярных каскадов экспрессии генов, обеспечивающей долговременное хранение информации при обучении и развитии. В процессе развития новые экстраклеточные сигналы ведут к инициации экспрессии генов, определяющей клеточную дифференциацию и консолидацию генетически запрограммированных функциональных систем. При обучении новые комбинации экстраклеточных сигналов вызывают реэкс-прессию генов, бывших активными в развитии. Это приводит к модификации функциональных систем и ее сохранению в долговременной памяти (поК.В, Анохину, 1997).
репрессию генома. Это достигается тем, что БР транспортируется в ядро к генам с помощью специального транслокационного белка (ТЛБ), который содержит метку активированного участка мембраны. После экспрессии генов (гена, кодирующего белок соот-ветствущего рецептора, и гена, ответственного за белок-метку) и синтеза соответствующих белков с помощью мРНК белок вместе с ТЛБ транспортируется к тому участку мембраны, который был закодирован белком-меткой. Эта схема предполагает, что механизм
экспрессии генов используется избирательно для изменения проводимости через селективные синапсы.
Большие возможности для выяснения молекулярных механизмов памяти открывает изучение собственно модификаций генома, вызываемых обучением. Исследование изменений ДНК под влиянием обучения в значительной степени тормозилось общепринятым представлением о «неприкосновенности» ДНК и ее причастности только к хранению генетической (врожденной) информации- В течение многих лет в молекулярной генетике господствовала идея о постоянстве генома. Однако в процессе жизнедеятельности клетки молекулы ДНК не остаются неизменными: отдельные участки ДНК умножаются, выпадают, перемещаются и модифицируются. Изменилось представление молекулярных биологов и о процессах транскрипции и трансляции. Мнение о том, что считывание генетической информации с ДНК всегда идет только в одном направлении; ДНК—>РНК—>белок, подверглось критике.
В 70-х годах Д. Балтимор (ВаШтог О.) и X. Темин и С. Мицута-ни (Тепип Н., М1гийт 5.) одновременно в одном и том же журнале «г^аШге» опубликовали данные о существовании фермента — обратной транскриптазы (или ревертазы), который способствует синтезу ДНК на основе информации, содержащейся в РНК, Так было открыто явление обратной транскрипции— передача информации в обратном направлении, от РНК к ДНК.
Изучение активности процесса обратной транскрипции при выработке у крыс пищедобывательного двигательного рефлекса в зависимости от их способности к обучению показало, что у быстро обучающихся животных активность обратной транскрипции в гиппокампе, которая измерялась по РНК-зависимому синтезу ДНК, в два раза выше по сравнению с медленно обучающимися животными (Р.И. Салганик и др., 1981). Группы животных составлялись селекционным методом. В одной наследственно закреплялась способность к быстрому обучению, другая формировалась из «неспособных:» крыс, медленно обучающихся животных.
Открытие обратной транскрипции (получение ДНК с копий РНК) позволяет предполагать, что существует процесс интеграции новых ДНК в геном, что может обеспечивать усиленный синтез клеточных структур, необходимых для сохранения новых связей между нейронами.
Экспериментальные доказательства модификации генома при обучении получены Н.А. Тушмаловой и ее сотрудниками (1973, 1993). Возрастание функциональной активности генома при обучении оценивалось двумя методами: по степени метилирования ДНК мозга и избирательной индукции синтеза ДНК. М-етилирова-
ние относится к обратимой модификации ДНК, т.е. не передаваемой по наследству. Изменение степени метилирования ДНК исследовалось у крыс под влиянием выработки трех групп условных рефлексов: пищевых, пищедобывательных (чтобы получить пищу, крыса нажимала на полочку-педаль) и оборонительных — при электрокожном подкреплении. Измерения производились в коре, гип-покампе и мозжечке. Контролем служили животные, получавшие условные и безусловные раздражители в случайном порядке, и животные, не участвующие в опытах с обучением. Исследователи обнаружили увеличение степени метилирования ДНК при выработке и угасании условных рефлексов. Максимальное увеличение метилирования соответствует ранним стадиям обучения, с закреплением условного рефлекса оно возвращается к исходному уровню. Сильнее оно представлено в гиппокампе, чем в коре, и в нейронах, чем в глии.
Повышение степени метилирования в процессе обучения свидетельствует об ускорении транскрипции, так как метилирование (присоединение метильных групп СН^) рассматривается как «метка» отрезков ДНК, с которых осуществляется транскрипция. Метилирование — один из молекулярных механизмов экспрессии генома.
Модификация генома под влиянием обучения была продемонстрирована также по показателю индукции синтеза ДНК. Синтез носил строго избирательный характер, он затрагивал главным образом последовательности ДНК, повторенные в геноме от 10 до 20 раз (ДНК умеренных или средних повторов) (Тушмалова Н.А., 1993; Ашапкин В.В. и др., 1983). Существует гипотеза, что на умеренных повторах ДНК идет активный синтез РНК и что повторы расположены перед геномом и являются сигналами, регулирующими транскрипцию. Н.А. Тушмалова предполагает, что избирательно индуцированный на определенных фрагментах синтез ДНК ответствен за активное накопление информации на ранней стадии обучения.
Анализ нуклеидного состава вновь синтезированной ДНК в неокортексе крысы, однако, показал, что индуцированная обучением ДНК существенно не отличается от суммарной. Поэтому индукция синтеза ДНК, наблюдаемая при обучении, по-видимому, связана с экспрессией генов, определяющих усиление процессов репарации ДНК. Продемонстрировано увеличение синтеза ДНК в коре головного мозга крысы под влиянием психотропного препарата (пирацетама), который применяется в клинике для лечения больных, страдающих нарушением памяти.
Недостаток многих гипотез о молекулярных механизмах памяти состоит в том, что авторы не учитывают наличие в клетках
эукариотов не одного, а многих типов ДНК. В ядре клетки имеется ядерная ДНК, включающая генетическую ДНК, сателлитная ДНК и ДНК ядерной оболочки. В платоплазме выделяют цитоплазмати-ческую (свободно плавающую) ДНК и ДНК клеточных органелл, среди них ДНК митохондрий (Тушмалова Н.А., 1990). Из всех типов ДНК наиболее изучена ядерная, генетическая ДНК.
По гипотезе Н.А. Тушмаловой, функция хранения генетической информации закреплена за стабильной (высокомолекулярной) ядерной ДНК. Кодирование же приобретенной информации выполняется лабильной (низко молекулярной) сателлитной ДНК ядра, которую характеризует непрочная связь с белком. Кроме того, для сателлитной ДНК как кандидата на хранение приобретенной памяти характерно высокое содержание метилированных групп, число которых особенно возрастает с обучением.
В своей гипотезе о параллельном кодировании врожденной и приобретенной памяти на разных типах ДНК (стабильной и лабильной) Н.А. Тушмалова основывается на сходстве некоторых биохимических реакций в онто- и филогенезе; экспериментальных данных о химической гетерогенности ДНК; существовании обратной связи в триаде ДНК — РНК — белок (обратной транскрипции).
Успешность обучения во многом зависит от сохранности операций, совершаемых на клеточном уровне, которые кодируются в генетической ДНК. Генетические исследования на плодовой мушке Ого50рЫ1а показали зависимость их обучения от синтеза адени-латциклазы. Обнаружены моногенные мутанты ОговорЬИа, у которых нарушено обучение. Оно тестировалось по выработке условного оборонительного рефлекса избегания специфического запаха после его подкрепления ударом тока. У одного такого мутанта имелся дефектный ген — гШаЬа^а, кодирующий аденилатциклазу, зависимую от комплекса кальция с калъмодулином. В результате такой мутации аденилатциклаза утратила способность активироваться комплексом кальция с кальмодулином. У ОгозорЬПа аденилатциклазой богаты особые отделы мозга (грибовидные тела), имеющие важное значение для некоторых типов ассоциативного обучения. У молодых мушек в грибовидном теле быстро растет число новых нейронов, что связывают с накоплением опыта в процессе обучения. Хирургическое разрушение грибовидного тела у древесных муравьев нарушает основы их «социального поведения». Другой мутант ОгозорпИа — аипсе («тупицы») обучается, но обладает поразительно короткой памятью в несколько десятков секунд. Оказалось, что мутация затронула у него фосфодиэстеразу, расщепляющую цАМФ, что приводило к увеличению содержания этого вещества. Видимо,
11-3341
слишком большие и слишком малые концентрации цАМФ препятствуют закреплению следов памяти. У всех мутантов с нарушением ассоциативного обучения нарушен и процесс сенситизации. Очевидно, что существует большое сходство механизмов пластичности у Ого50рЫ1а и у моллюска, они одинаково используют ней-ромедиатор, выполняющий модулирующую функцию, и фосфо-рилирование белков для получения более стойкого эффекта.
Способность мясной мухи и ОговорпПа к обучению наследуется, что доказывается опытами со скрещиванием. Для выведения особей, способных и неспособных к обучению, для скрещивания соответственно отбирали из быстро и медленно обучающихся. Отбор способных мух к выработке условного рефлекса на сладком подкреплении (втягивание хоботком сладкой воды) показал, что только к 24-му поколению прекращается рост кривой эффективности выработки условного рефлекса по мере выведения все более способных к обучению особей. В связи с тем что процесс роста эффективности обучения идет медленно — только к 24-му поколению, — говорят не об одном, а о многих генах, определяющих обучаемость. В то же время формирование пластичности по типу сенситизации у мясной мухи определяется одним геном. Кривая эффективности обучения выходит на плато уже через одно поколение.
Таким образом, успешность обучения зависит от сохранности набора генов, опосредующих обучение, а ген сенситизации — один из участников генетического пула, управляющего обучением.
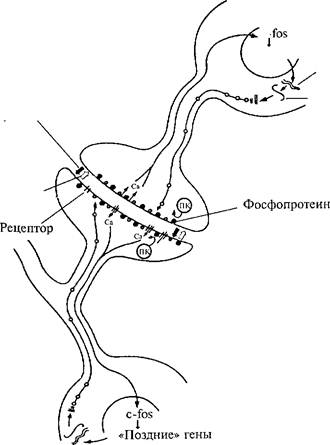
Все биохимические и молекулярные преобразования, вызываемые обучением, в конечном счете ведут к синаптической реорганизации — увеличению размеров, количества активно работающих синапсов. Существуют две гипотезы относительно изменений синаптических контактов под влиянием обучения: функциональная и структурная. Согласно функциональной теории обучение не изменяет число синапсов. Оно остается постоянным, но при этом увеличивается отношение активных синапсов к пассивным. По структурной гипотезе обучение стимулирует образование новых контактов между нейронами, которые не берутся из резерва, а создаются заново-Основные модификации, наблюдаемые в процессе обучения на клеточном уровне и ведущие к структурным преобразованиям контактов между нейронами, по результатам исследования С. Роу- за (1995) отражены на рис. 38. Изучая обучение у цыплят (избегание клевания горькой бусинки), он выделил несколько уровней преобразований в клетке, которые сохраняются в течение разных отрезков времени ~ на разных этапах жизни энграммы.
| Глутамат |
| С. Г05 «Поздние» гены РНК |
| Белок |
| Гликопротеин |
| «Ретроградный» сигнал |
|
Рис. 38. Основные молекулярные и структурные модификации нейрона, возникающие в процессе обучения.
Показан каскад биохимических реакций в синапсе и ядре в процессе формирования памяти (по С. Роузу, 1995).
В течение секунд и минут сохраняются изменения, вызываемые стимулом в ММОА-рецепторах, отвечающих на глутамат, а также в мускариновых рецепторах, чувствительных к АХ. Сохранение следового эффекта в течение минут или часов определяется действием киназ, которые фосфорилируют пресинаптические белки ионных каналов, а также последующей экспрессией ранних генов (с-Сок, с-]ип). Время сохранения памяти от 1 до 6 ч связывают с синтезом новых белков — глюкопротеинов в результате экспрес-
1Т
сии поздних генов. Глюкопротеины создают структурные изменения, встраиваясь в пресинаптические и постсинаптические участки мембран. Период 12—24 ч связан с образованием дополнительных шипиков, увеличением их диаметра и площади постсинапти-ческих уплотнений, которые выявляются с помощью электронного микроскопа. Постсинаптические уплотнения образованы плотно пригнанными рецепторами.
Гл а в а 7 ЭМОЦИИ
|
|
|
|
|
Дата добавления: 2014-12-10; Просмотров: 684; Нарушение авторских прав?; Мы поможем в написании вашей работы!