КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Вегетативное, бесполое и половое размножение водорослей. Циклы развития водорослей. Изоморфное и гетероморфное чередование поколений
|
|
|
|
Рост бактериальной клетки и популяции. Периодические культуры. Фазы кривой роста периодической культуры. Сбалансированный и несбалансированный рост, причины лимитации роста и отмирания. Проточные и синхронные культуры.
Рост прокариотич.кл-соглассованное увелич-ние всех химич. Комп-тов, их кот. Она построена. Рост явл-ся рез-том множества скоординированных Биос-ких пр-сов, наход-ся под строгим регул-м контролем и приводит к увеличению биомассы и размеров клетки. При достижении определ-х размеров рост прекращàкл делится. Рост нового мат-ла различен. У кокков-рост по всей пов-ти, У E. colli в виде локальных зон роста, которые распол. В местах адгезии цитопл. Мембраны и внешней мембраны этих м/о. В центре формир-ся септа. Кл-ки, возникшие из 1 материнской-новая генерация. Время генерации- время от возникновения до деления Кл-ки. Чем < организм, тем < время генерации. E. Colli время генерации 20 минут. В оптимальных условиях сущ-т мех-мы регуляции числ-ти, ограничения роста, развития м/о. Проследить динамику роста бактериальной поп-ции можно на периодической к-ре-культуре, выращ-ой в лабораторных условиях без притока свежей среды и оттока метаболитьв. В таких условиях рост бактер. Поп-ции имеет несколько стадий:1) Лагфаза -подготовительная. Наблюдается при внесении Кл-к в свежую среду. В теч-нии лагфазы-индукция ферм-тов для расщепл-я новых субстратов. Длит-ть зав от состава новой среды (> разница м/у новой и старойàдольше подготовительный этап); V внесенного мат-ла (> мат-лаà< подготовит. Этап);качества и возраста к-ры (чем старше, тем >).2) Логфаза - фаза экспоненциального роста. Наблюдается логорифмическая зав-ть числа Кл-ток от времени выращивания: μ=lnx1*lnx0/t1-t0. Фаза хар-ся постоянной скоростью роста и деления клеток, дл-ть зависит от штамма бактерий и питат. Среды. Вел-на Кл-то и состава макромол-л не меняется (кол-во белка, ДНК, РНК постоянно). Такой рост называется сбалансированным- кол-во С и Е субстрата распределено пропорционально м/у различными комп-тами Кл-к. Сод-ние РНК > по сравн. С ДНК и белком, т.к. скорость синтеза зависит от числа рибосом. Во время перехода к стационарной фазе рост становится несбалансированным. При возрастании биомассы накапл. Пр-ты обмена, исчерпыв-ся источники питания, закисл. Среда, сниж-ся кислородàнеравномерное распред-ние C и Е м/у комп-ми Кл-киà состав Кл-ки меняется. В условиях несбал. Роста пр-дит запас пит. В-в(полиβ-гидроксибутират, гликоген и др.), м.б. отравление пр-ми метаб-маà торм-ние биосинтетич. Пр-сов, потребл-ние питат. Субстратов продолжается. Сод-ние РНК снижается.3) Стационарная фаза -изм-ние числ-ти кл не пр-дит. Отмирание =скорости размножения. Кол-во биомассы в стац. Фазу- урожай. Кол-во урожая зав от состава пит. В-в, условий культив-ния, вида м/о;4) Фаза отмирания - отмирающая культура> чем размнож. Отмирание тоже носит экспоненциальный х-р. Причины гибели: рез-тат совместного действия множества ф-ров-исчерпание С, О, накопл-ние пр-тов метаболизма, воспр-я кворума- воспр. Ацетата. Для E. Colli на синтетич. Минер. Средах накопл-е ацетата-рез-та сбраживания пит. Субстратовàотмирание м/о. Накопл-е ацетата-проблема генной инженерии. Кривая роста -модель. Отражающая рост периодич. Культуры, была предложена Ж. Мано. Проточные к-ры:культуры, выращенные в спец. Культиваторах. Исп-т в промышл-х целях для получения большой биомассы. Исп-т хемо-и-турбидостаты-меняют режим регуляции. Хемостат-культиватор-сосуд с мешалкой. Постоянная скорость перемешиванияàравномерная аэрация. В культиватор подается свежая среда и удаляется часть к-ры. При пост-нии свежей среды-разбавление м/о в культиваторе. Кл-ки интенсивно размножаются за сч. Субстрата. Разбавл-ние уравнивается скоростью деления клеток. Основное условие хемостата-1 из комп-тов пит. Среды в лимитирующей к-цииàфизиол. Состояние культуры в культиваторе-переход к стац. Фазе роста. Турбидостат-комп-ы пит. Среды в избыткеà эксп. Фаза роста. Рег-я числ-ти Кл-к на основе контроля мутности в культиваторе. Устан. Датчики оптич. Плотности. Если фактическая плотность изм. От заданной, автоматич. Изменяется скорость подачи среды в культиватор, плотность поддержив-ся на пост. Уровне. Синхронные к-ры: к-ры, где дел-е Кл происходит одновременно. Способы достиж-я синхронного деления-выдерживание при низких темп-рах;отбор по размеру(посде дел-я Кл миним разме, до дел-максим.;центрифугир-е в градиенте плотности(мелкие-в верхн. Части, крупные-внизу); фильтрация-быктер. Фильтры с заданным размером пор; вынужденное гол-ниеàмол. Кл. стареют.
|
|
|
|
|
|
Вегетативное размножение у одноклеточных водорослей происходит делением клеток. У колониальных – распадением колоний или образованием новых колоний внутри клеток материнский особи (у вольвоксовых и многих протококковых). У нитчатых водоро слей – случайным разрывом нитей или распадением - фрагментация (у сине-зеленых, у зигнемовых). У немногих водорослей образуются специальные органы вегетативного размножения – клебеньки (у харовых).
Бесполое размножение широко распространено у водорослей и осуществляется образованием специальных спор, покидающих материнскую клетку. У большинства эти споры подвижны, имеют монадную структуру, неодеты оболочкой – зооспоры, образуются несколько, реже по одной в зооспорангиях. Они не отличаются от вегетативных клеток. Зооспоры, поплавав непродолжительное время, одеваются оболочкой и прорастают в новую водоросль. У других водорослей споры бесполого размножения неподвижны – апланоспоры (автоспоры).
Половое размножение. У немногих НВ (вольвокс) сливаются целиком две особи, наиболее примитивный тип полового процесса – хологамия. У громадного большинства водорослей половой процесс состоит в слиянии (копуляция) двух голых половых клеток (гамет) в одну клетку (зигота). В зиготе (как и при хологамии) происходит слияние протоплазм обеих гамет и их ядер. Зигота покрывается оболочкой, нередко очень толстой и/или немедленно прорастает (у морских водорослей), или, главным образом у пресноводных водорослей, переходит в состояние покоя, после чего прорастает или образует зооспоры, или растение.
|
|
|
Если обе сливающиеся гаметы подвижны и одинаковы по форме и величине, их называют изогаметами, а половой процесс изогамией. Если обе гаметы подвижны одинакого, но одна значительно больше другой, их называют гетерогаметами, а ПП гетерогамия. Более крупная гамета считается женской, меньшая – мужской. Клетки, в которых образуются изо - или гетерогаметы называют гаметангиями. Если копулируют друг с другом гаметы, развивавшиеся на одной особи – гомоталличные, если же, развивавшиеся на разных особях – гетероталличные. Внешне морфологических отличий у гетероталличных особей разных полов не бывает, и если их гаметы не различимы (изогаметы), то такие особи различающиеся лишь физиологически по полу, обозначают как плюс и минус особи.
Высшим типом полового процесса у водорослей является оогамия; мужская подвижная маленькая гамета – сперматозоид, который оплодотворяет более крупную неподвижную яйцеклетку. Клетку, в которой формируются сперматозоиды называют антеридием; а клетку, содержащую яйцеклетку (одну, реже несколько)- оогонием.
Имеется ещё особый тип полового процесса – зигогамия (спирогира), как конъюгация, при которой две клетки соединяются поперечными выростами и сливаются их протопласты, не имеющие жгутиков и не дифференцированные внешне на мужскую и женскую гаметы.
Слияние клеточных ядер при половом процессе влечет за собой увеличение в получающемся ядре зиготы числа хромосом вдвое. Это удвоенное число хромосом называется диплоидным, а вдвое меньшее число хромосом – гаплоидным. У НР и частности у водорослей, редукционное деление в разных систематических группах происходит в различное время их жизненного цикла. У одних имеется правильное чередование бесполого поколения - спорофита и полового – гаметофита и редукционное деление происходит, как у ВР перед образованием на спорофите спор, из которых развивается гаметофит. У очень многих водорослей первое же деление ядра в зиготе редукционное и весь жизненный цикл происходит в гаплоидной фазе; диплоидная лишь зигота, которую трудно считать «поколением», и у таких водорослей говорят о смене ядерных фаз. Одна лишь смена ядерных фаз без чередования поколений имеет место и в случаях, когда жизненный цикл всей водоросли происходит в диплоидной фазе, а в гаплоидной фазе лишь гаметы, перед образованием которых происходит редукционное деление. Если нет митотического деления, это приводит к неодинаковому временному соотношению гаплоидной и диплоидной стадии. В этом случае зигота имеет мейотическое деление (зиготическая редукция хромосом). Получающиеся из зиготы растения гаплоидны. Эта стадия у большинства продолжительная. Гаплоидная стадия будет и при бесполом размножении у зоо - и автоспор. При зиготической редукции хромосом доплоидны только зиготы, а все остальные стадии гаплоидны. Диплоидная стадия начинается с слияния гамет и заканчивается мейозом зиготы. Зигота претерпевает редукционное деление, потому развивающиеся из нее растения гаплоидные. На гаплоидных растениях могут формироваться зоо- и автоспоры, но могут прорастать на растения гаметы гаплоидные. Гаметы, сливаясь, дают диплоидную зиготу.
|
|
|
|
 1) О=>R'®Оn®зооспоры (n)®On
1) О=>R'®Оn®зооспоры (n)®On
 2n гаметы (n)
2n гаметы (n)
 гаметы (n)
гаметы (n)
гаметы (n)
2) О®О®зооспоры (2n)
 2n
2n
 |
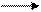 R®гаметы
R®гаметы
 |
2n
зооспоры
Спорическая редукция хромосом: наблюдается при образовании зооспор. Гаплоидные и диплоидные стадии равновелики (морской салат – Ulva). Эти растения называются спорофиты. Из гаплоидных зооспор вырастают растения, внешне похожи на диплоидные.
3)
|

 | |||||||||
 | |||||||||
 | |||||||||
 | |||||||||
| |||||||||
 | |||||||||
|
 только зооспоры
только зооспоры 
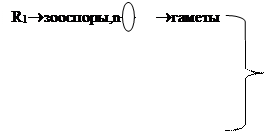
 R1®зооспоры,n®гаметофит ®гаметы
R1®зооспоры,n®гаметофит ®гаметы
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 840; Нарушение авторских прав?; Мы поможем в написании вашей работы!