КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Клітинні і неклітинні форми життя 1 страница
|
|
|
|
Основними формами життя на Землі є організми клітинної будови. Цей тип організації характерний для всіх видів живих істот, за винятком вірусів, які розглядають як неклітинні форми життя. Вони настільки малі, що лише в кілька разів перевищують розміри великих молекул білків. Віруси мають розміри 10—275 нм. їх можна побачити лише під електронним мікроскопом. Вони легко проходять крізь пори спеціальних фільтрів, що затримують усі бактерії і клітини багатоклітинних організмів. Віруси були відкриті у 1892 р. російським фізіологом рослин і мікробіологом Д. І. Іванівським під час вивчення хвороби тютюну.
Віруси — збудники багатьох хвороб рослин і тварин. Вірусними хворобами людини є кір, грип, гепатит А (хвороба Боткіна), поліомієліт (дитячий параліч), сказ, віспа тощо.
Під електронним мікроскопом різні види вірусів мають форму паличок або кульок. Окрема вірусна часточка складається з молекули нуклеїнової кислоти (ДНК або РНК), скрученої в клубок, і молекул білка, розміщених у вигляді своєрідної оболонки навколо молекули кислоти (капсид). Віруси не здатні самостійно синтезувати нуклеїнові кислоти і білки, з яких вони складаються. Розмноження вірусів можливе лише в разі використання ферментних систем клітин. Потрапивши у клітину, віруси змінюють і перебудовують обмін речовин у ній, внаслідок чого клітина починає синтезувати молекули нових вірусних часточок. Поза клітинами віруси переходять у кристалічний стан, що сприяє їх збереженню.
У житті вірусів можна виділити такі етапи: прикріплення вірусу до клітини, вторгнення вірусу в клітину, латентну стадію, утворення нового покоління вірусів, вихід вібріонів. У латентну стадію вірус ніби зникає. Його не вдається виявити або виділити з клітини, але в цей період уся клітина синтезує необхідні для вірусу білки і нуклеїнові кислоти, в результаті чого утворюється нове покоління вібріонів.
Проникнення вірусу в клітину організму хазяїна розпочинається із взаємодії вірусної часточки з поверхнею клітини, на якій є особливі рецепторні ділянки. Оболонка часточки вірусу має відповідні прикріпні білки, які "впізнають" ці ділянки. Саме цим зумовлена висока специфічність вірусів стосовно клітин-хазяїв: часто віруси уражують лише певний тип клітин якогось виду організмів. Так, вірус поліомієліту уражує лише нервові клітини людини, а вірус тютюнової мозаїки — клітини листків тютюну. Якщо часточка вірусу прикріплюється не до рецепторних ділянок, а до інших місць на поверхні клітини-хазяїна, то зараження останньої може і не відбутися. Отже, наявність рецепторних ділянок на поверхні клітини визначає її чутливість до того чи іншого виду вірусів.
Усередину клітини-хазяїна вірус може проникнути різними шляхами. Часом оболонки вірусних часточок зливаються з клітинною мембраною (як у вірусу грипу), і ДНК виявляється у цитоплазмі клітини, іноді вірусна часточка потрапляє в клітину шляхом піноцитозу, після чого ферменти клітини-хазяїна розщеплюють її оболонку і вивільняють нуклеїнову кислоту (вірус поліомієліту тварин). У рослинні клітини віруси можуть проникати крізь пошкоджені ділянки клітинної стінки.
У 1917 р. французький вчений Ф. д'Ерелл відкрив віруси бактерій — бактеріофаги (або фаги). Під електронним мікроскопом вони мають форму коми або тенісної ракетки розміром близько 5 нм. Коли часточка фата прикріплюється своїм тонким відростком до бактеріальної клітини, його ДНК проникає в клітину і викликає синтез нових молекул ДНК і білка бактеріофага. Через ЗО—60 хв бактеріальна клітина руйнується і з неї виходять сотні нових часточок фага, здатних спричинити зараження інших бактеріальних клітин.
Спочатку вважали, що бактеріофаги можна використовувати для боротьби з хвороботворними бактеріями. Проте згодом виявилося, що фаги швидко руйнують бактерії в пробірці, але неефективні в живому організмі. У зв'язку з цим їх використовують в основному для діагностики захворювань, виявлення бактерій.
Клітинна теорія. Відкриття і вивчення клітини стало можливим завдяки винайденню мікроскопа та вдосконаленню методів мікроскопічних досліджень. Перший опис клітини зроблено у 1665 р. англійським ученим Р. Гуком. Згодом з'ясувалося, що він відкрив не клітини (у власному розумінні терміна), а лише зовнішні оболонки рослинних клітин.
Прогрес у вивченні клітини пов'язаний з розвитком мікроскопії у XIX ст. На той час уявлення про будову клітини змінилось: за основу клітини бралася не клітинна оболонка, а її вміст — протоплазма. У протоплазмі було відкрито постійний компонент клітини — ядро. Численні відомості про тонку будову та розвиток тканин і клітин дали змогу зробити узагальнення. Таке узагальнення запропонував у 1839 р. німецький біолог Т. Шванн у вигляді сформульованої ним клітинної теорії. Він стверджував, що клітини рослин і тварин принципово подібні між собою (мал. 1).
Крім Т. Шванна та М. Шлейдена, співавторами клітинної теорії вважають німецького вченого Рудольфа Вірхова та естонського вченого Карла Бера. Р. Вірхов довів, що клітини утворюються з інших клітин внаслідок їх поділу. К. Бер відкрив яйцеклітину птахів і ссавців і довів, що
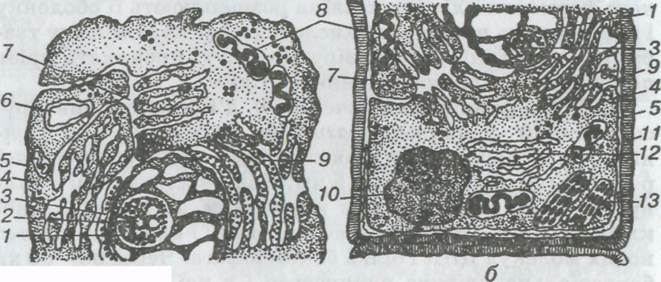
Мал. 1. Будова тваринної (а) і рослинної (б) клітин:
1 — ядро; 2 — ядерна мембрана; 3 — ядерце; 4 — протоплазма; 5 — плазматична мембрана; 6 — лізосома; 7 — комплекс Гольджі; 8 — мітохондрії; 9,12 — відповідно шорстка і гладенька ендоплазматична сітка; 10 — вакуоля; 11 — клітинна оболонка; 1З — хлоропласт
багатоклітинні організми цих тварин розвиваються з однієї клітини — заплідненої яйцеклітини (зиготи). Отже, клітина є одиницею не лише будови, а й розвитку організму.
Створення клітинної теорії стало найважливішою подією в біології, одним із вирішальних доказів єдності всієї живої природи. Клітинна теорія сприяла розвитку ембріології, гістології і фізіології. Вона стала основою для матеріалістичного розуміння життя, пояснення еволюційного взаємозв'язку організмів, розуміння суті індивідуального розвитку.
Основні положення клітинної теорії зберегли своє значення і нині, хоча більш ніж за 100 років були отримані нові відомості про структуру, життєдіяльність і розвиток клітин.
Сучасна клітинна теорія включає такі положення:
• клітина — елементарна одиниця будови і розвитку всіх живих організмів;
• клітини всіх одно- і багатоклітинних організмів подібні за походженням (гомологічні), будовою, хімічним складом, основними виявами життєдіяльності;
• кожна нова клітина утворюється тільки внаслідок розмноження материнської клітини шляхом поділу;
• у багатоклітинних організмів, які розвиваються з однієї клітини (зиготи, спори тощо), різні типи клітин формуються внаслідок їхньої спеціалізації упродовж індивідуального розвитку особини й утворюють тканини;
• з тканин складаються органи, які тісно пов'язані між собою й підпорядковані нервово-гуморальним та імунним системам регуляції.
Клітинна теорія є одним з найбільших узагальнень природознавства XIX ст. Формування клітинної теорії створило основу для подальших фундаментальних досліджень процесів життєдіяльності, будови та розвитку всіх організмів на Землі.
БУДОВА І ФУНКЦІЇ КЛІТИНИ
Наука, що вивчає будову, хімічний склад, процеси життєдіяльності і розмноження клітин, називається цитологією (гр. куtos — порожнина, 1оgos — наука).
Клітина — це структурна одиниця живих організмів, що є певним чином диференційованою ділянкою цитоплазми, оточеною клітинною мембраною. Функціонально клітина є основною одиницею життєдіяльності організмів.
Клітини існують і як самостійні організми, і входять до складу багатоклітинних організмів. Бактерії, багато видів водоростей {хлорела, хламідомонада), нижчих грибів (цукор, дріжджі) і найпростіші тварини (амеба, евглена зелена, інфузорії тощо) складаються з однієї клітини. Ця клітина виконує всі функції живого організму — живлення, руху, розмноження тощо. Тіло більшості видів рослин і тварин складається з величезної кількості клітин, які спеціалізуються на виконанні окремих функцій. Ці клітини утворюють різні тканини (тканини рослин — див. "Ботаніка", тканини тварин — див. "Анатомія, фізіологія і гігієна людини").
Хоча клітини (рослин, тварин і грибів) мають різну будову і виконують різні функції, вони мають багато спільних морфологічних особливостей (сформоване ядро, подібний набір органел) і подібних функціональних властивостей (біосинтез білків, використання і перетворення енергії, процеси розмноження).
Клітини різняться за розмірами, формою (мал. 2), особливостями організації, функціями. За формою клітини бувають: циліндричні і кубічні (епітеліальні тканини), дископодібні (еритроцити), кулясті (яйцеклітини), видовжені і веретеноподібні (м'язові), зірчасті (нервові) тощо. Серед клітин трапляються і такі, що не мають сталої форми. Це так звані амебоїдні клітини (наприклад, лейкоцити).
Більшість клітин багатоклітинного організму мають розміри від 10 до 100 мкм, а найдрібніші — 2—4 мкм. Великі розміри мають деякі рослинні клітини з великими вакуолями в цитоплазмі: клітини м'якоті кавуна, лимона (їх можна бачити неозброєним оком). Дуже великі розміри (до кількох сантиметрів у діаметрі) мають яйцеклітини птахів і деяких риб. Розміри клітин тваринних організмів не залежать від розмірів їхнього тіла. Наприклад, клітини печінки коня і миші мають майже однакові розміри. Відростки нервових клітин іноді досягають одного і більше метрів. Кількість клітин в організмі зазвичай дуже велика. Лише окремі багатоклітинні організми мають невелику кількість клітин. Наприклад, такі порівняно великі організми, як коловертки, складаються лише з 400 клітин. У хребетних тварин і людини найчисленнішими є клітини крові і головного мозку. Так, у людини кора мозку складається з (14...15) * 109 клітин, а клітин нейроглії у 5—10 разів більше. Кількість еритроцитів у крові дорослої людини становить 23 • 1012. Невеликі розміри і значна кількість клітин створюють у багатоклітинних організмів величезну поверхню, що важливо для забезпечення швидкого обміну речовин.
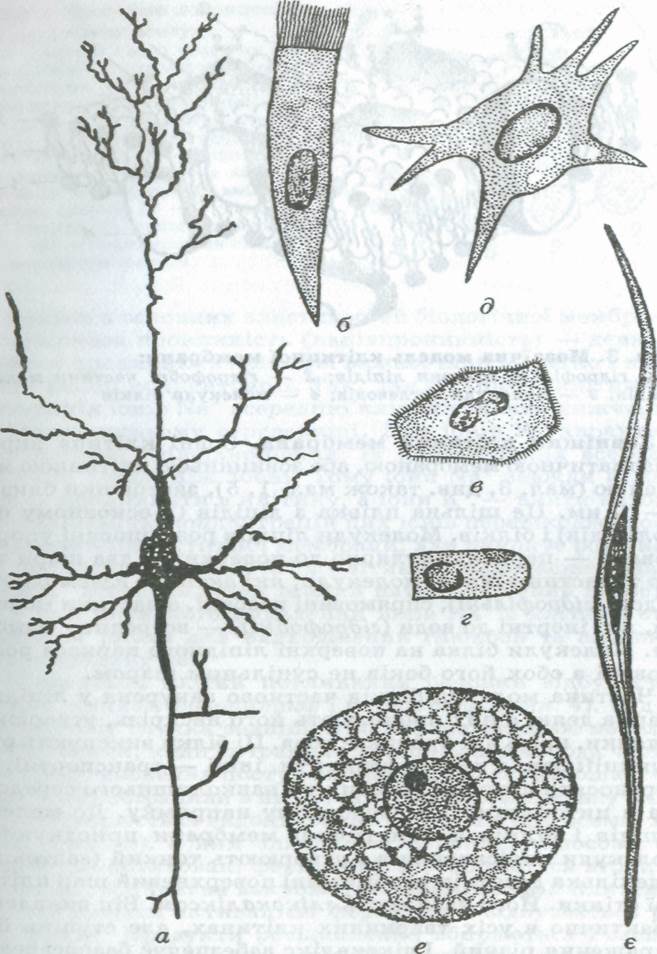
Мал. 2. Форма клітин організмів людини і тварин:
a — нервова клітина (нейрон); б—г — різні види епітеліальних клітин;
д— клітина сполучної тканини; е — яйцеклітина; є — м'язова клітина
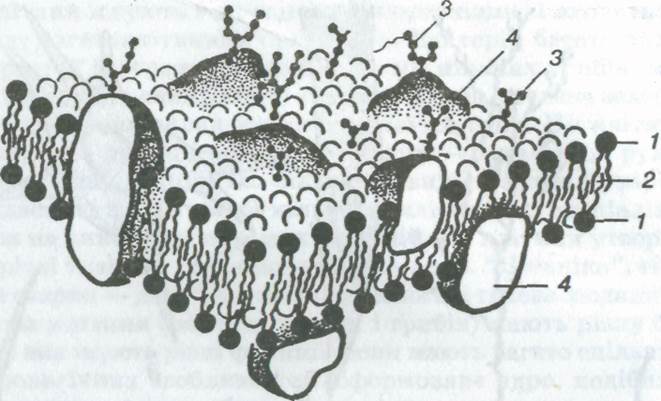
Мал. 3. Мозаїчна модель клітинної мембрани:
1 — гідрофільні головки ліпідів; 2 — гідрофобні частини молекул ліпідів; 3 — молекули вуглеводів; 4 — молекули білків
Зовнішня клітинна мембрана. Зовні клітина вкрита плазматичною мембраною, або зовнішньою клітинною мембраною (мал. З, див. також мал. 1, 5), завтовшки близько 6—10 нм. Це щільна плівка з ліпідів (в основному фосфоліпідів) і білків. Молекули ліпідів розташовані упорядковано — перпендикулярно до поверхні, у два шари так, що ті частини їхньої молекули, які активно взаємодіють з водою (гідрофільні), спрямовані назовні, а частини молекули, які інертні до води (гідрофобні), — всередину мембрани. Молекули білка на поверхні ліпідного каркаса розташовані з обох його боків не суцільним шаром.
Частина молекул білків частково занурена у ліпідний шар, а деякі з них пронизують його наскрізь, утворюючи ділянки, крізь які проникає вода. Ці білки виконують різні функції: одні з них — ферменти, інші — транспортні, які переносять потрібні речовини з навколишнього середовища в цитоплазму і у зворотному напрямку. До молекул ліпідів і білків плазматичної мембрани приєднуються молекули полісахаридів і утворюють тонкий (завтовшки у декілька десятків нанометрів) поверхневий шар клітинної стінки. Його називають глікокаліксом. Він виявлений практично в усіх тваринних клітинах, але ступінь його вираження різний. Глікокалікс забезпечує безпосередній зв'язок клітини із зовнішнім середовищем. У ньому відбувається позаклітинне травлення і розміщені клітинні рецептори.
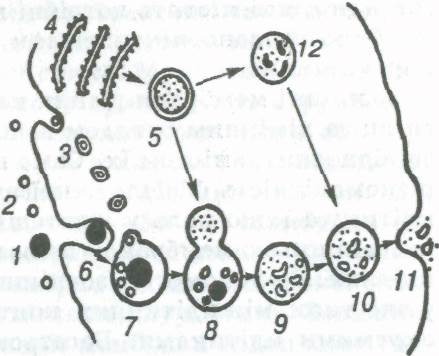
Мал. 4. Лізосомн в процесі фаго- і піноцнтозу:
1,2 — великі і малі часточки в середовищі, яке оточує клітину; захоплення часточок шляхом ліноиитозу (3, 7) і фагоцитозу (6, 7); 4, 5 — утворення лізосо'м (4 — гранулярна ендоплазматична сітка); 8 — виливання ферментів лізосом у вакуолі; 9,10 — розщеплення вмісту вакуолі (лізис); 11 — виділення продуктів розщеплення;12—ділянка цитоплазми після
дії ферментів лізосом.
Однією з основних властивостей біологічної мембрани є її вибіркова проникність (напівпроникність) — деякі речовини проникають крізь неї важко, інші — легко і навіть у бік вищої концентрації. Так, для більшості клітин концентрація іонів Nа+ всередині клітини значно нижча, ніж у навколишньому середовищі. Для іонів К+ характерне протилежне співвідношення: їх концентрація всередині клітини вища, ніж зовні. Тому іони Nа+ завжди намагаються проникнути в клітину, а іони К+ — вийти назовні. Вирівнюванню концентрацій цих іонів перешкоджає особлива система клітинної мембрани, яка виконує роль насоса, що відкачує іони Иа+ з клітини й одночасно накачує іони К+ всередину (так званий натрій-калієвий насос).
У разі активного видалення іонів Na+ з клітини створюються умови для надходження глюкози та амінокислот всередину неї.
У багатьох клітин поглинання речовин відбувається також шляхом фагоцитозу і піноцитозу (мал. 4). Під час фагоцитозу гнучка зовнішня мембрана утворює невеликі заглиблення, куди потрапляє захоплювана тверда часточка. Це заглиблення поступово збільшується, глибшає, часточки, які потрапили в нього, занурюються всередину клітини, й утворюється ендоцитозний пухирець (фагосома, харчова вакуоля). З ним зливається первинна лізосома (що містить неактивовані ферменти), й утворюється вторинна лізосома, або травна вакуоля (гетерофагосома). Злиття супроводжується активацією ферментів і відбувається розщеплення. Продукти розщеплення вбираються і засвоюються цитоплазмою клітини. Рештки виводяться назовні шляхом екзоцитозу. Явище фагоцитозу властиве амебам і деяким іншим найпростішим, а також лейкоцитам (фагоцитам). Аналогічно відбувається і поглинання клітинами рідин, які містять потрібні клітині речовини. Це явище було названо піноцитозом (гр. ріпо — п'ю, куіое — порожнина).
Зовнішні мембрани різних клітин істотно відрізняються як за хімічним складом власних ліпідів і білків, так і за відносним вмістом їх. Саме ці особливості визначають різноманітність фізіологічної активності мембран різних клітин та їхню роль у життєдіяльності клітин і тканин. Із зовнішньою мембраною зв'язана ендоплазматична сітка клітини. За допомогою зовнішніх мембран здійснюються різні типи міжклітинних контактів, тобто зв'язок між окремими клітинами. Багатьом типам клітин властива наявність на їхній поверхні великої кількості виступів, складок, мікро ворсинок. Вони значно збільшують площу поверхні, прискорюють обмін речовин та зміцнюють зв'язки окремих клітин між собою. У рослинних клітинах зовні клітинної мембрани є товсті оболонки, які добре видно в оптичний мікроскоп, що складаються з клітковини — целюлози (див. мал. 1, поз. 11). Ці вторинні клітинні стінки — продукт життєдіяльності органел цитоплазми (зокрема, комплексу Гольджі). Вони створюють міцну опору рослинним тканинам (деревина). Деякі клітини тваринних організмів також мають низку зовнішніх структур, що вкривають мембрану і виконують захисну функцію. Прикладом може бути хітин покривних клітин комах.
Товста целюлозна оболонка не повністю ізолює рослинні клітини одну від одної. У клітинах багатьох рослинних тканин ця оболонка пронизана численними канальцями, крізь які проходять вирости цитоплазми (плазмодесми), що сполучають сусідні клітини. Така структура забезпечує надходження продуктів життєдіяльності з однієї клітини в іншу. Цитоплазма. Внутрішній вміст клітини (протоплазма) ділиться на цитоплазму і ядро. Цитоплазма є основною за об'ємом частиною клітини. За фізичними властивостями це напіврідка маса колоїдної структури, в якій містяться органели клітини мембранної (ендоплазматична сітка, мітохондрії, пластиди, комплекс Гольджі, лізосоми) і не мембранної (рибосоми, центріолі клітинного центру) будови. Агрегатний стан цитоплазми може бути різним: рідким — золь і в'язким — гель. За хімічним складом цитоплазма досить складна.
Під оптичним мікроскопом у цитоплазмі не вдається розгледіти жодних структур, і вона здається однорідною. За допомогою електронного мікроскопа було встановлено, що в цитоплазмі всіх клітин тваринного і рослинного походження є складна система мембран, які часто розміщуються паралельно одна одній. Ця система мембран дістала назву ендоплазматичної сітки. Мембрани завтовшки
6—8 нм мають ліпідно-білкову природу і за структурою
подібні до зовнішньої мембрани клітини. Вони обмежують дуже розгалужену взаємозв'язану систему канальців, щілин і міхурців, яка сполучає різні ділянки клітини. Діаметр порожнини канальців — 25—30 нм. Мембрани ендоплазматичної сітки безпосередньо зв'язані з мембранами комплексу Гольджі. З мембран ендоплазматичної сітки утворюється оболонка ядра після поділу клітини. Зовнішня оболонка ядра є продовженням мембрани ендоплазматичної сітки і навколо ядерний простір сполучений з простором ендоплазматичної сітки.
Ендоплазматичні мембрани бувають двох типів: гладенькі (агранулярні) і шорсткі (гранулярні) (див. мал. 1, поз. 9, 12). Стінки останніх несуть на собі багато рибосом. Ендоплазматична сітка збільшує внутрішню поверхню клітини, що важливо для процесів обміну, бере активну участь у біосинтезі білків, жирів і вуглеводів, транспортує хімічні речовини в різні ділянки клітини.
Крім ендоплазматичної сітки в цитоплазмі є структури не мембранної будови — мікро трубочки. Це тоненькі ниткоподібні структури завтовшки 15—20 нм, до складу яких входять скоротливі білки. Вони утворюють нитки ахромати нового веретена під час мітозу, входять до складу клітинного центру, а також війок і джгутиків клітини. Вважають, що мікро трубочки мають значення для підтримання форми клітини (цитоскелет), а також забезпечують внутрішньоклітинний рух (пересування хромосом) і рух клітини загалом за допомогою джгутиків і війок.
Крім мікро трубочок до складу цитоскелета клітини еукаріотів входять мікрофіламенти та проміжні філаменти. Мікро філаменти складаються в основному з білка актину і трапляються в усіх клітинах еукаріотів. Вони входять до складу м'язових волокон та спеціальних клітинних компонентів (мікро ворсинок), утворюючи пучки в цитоплазмі рухливих клітин тварин. У багатьох рослинних клітинах і клітинах нижчих грибів вони розміщені в шарах рухливої цитоплазми. У клітині вони або є частиною скоротливого апарату, або беруть участь у формуванні жорстких скелетних структур, або утворюють жорстку фібрилярну сітку. Проміжні філаменти побудовані з фібрилярних білків. Це справжня опорна система клітин, що зазнають значних фізичних навантажень (епітеліальні клітини). В аксонах проміжні філаменти (нейрофіламенти) створюють жорстку основу, яка забезпечує гнучкість і цілісність тонких цитоплазматичних відростків нервових клітин. У смугастих м'язах проміжні філаменти входять до складу зет-дисків.
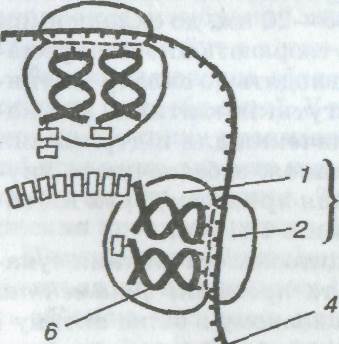
Рибосоми також було виявлено лише за допомогою електронного мікроскопа. Рибосома — органела сферичної форми діаметром 15—20 нм, що складається з молекул рибонуклеїнової кислоти (4 молекули: одна в малій і три у великій субодиницях у еукаріотів, у прокаріотів відповідно 1 і 2) і білка. Кожна рибосома має дві неоднакові субодиниці (субчастинки) — велику і малу (мал. 5), Форма і конфігурація рибосом різних організмів і клітин (прокаріотів і еукаріотів) напрочуд подібні, хоча й різняться в деталях. Утворюються рибосоми в ядерцях у вигляді окремих субодиниць. Субодиниці об'єднуються в рибосому поза ядром. Мала рибосомальна субодиниця має активну ділянку для приєднання матричної і транспортної РНК, а велика — для синтезу поліпептидного ланцюга. Частина рибосом сполучена з мембранами ендоплазматичної сітки (шорстка ЕПС), інша — розміщена в цитоплазмі. Основною функцією рибосом є матричний синтез білків, під час якого рибосоми зв'язують і утримують компоненти бі-
локсинтезуючої системи, виконують каталітичні функції і трансляцію. Особливо багато рибосом у клітинах тканин, які швидко ростуть.
Мітохондрії — орга-нели клітин, які добре видно під оптичним мікроскопом. Вони мають форму паличок, зерен, ниток розміром від 0,5 до 5 мкм. Мітохондрії нервових клітин мають довжину до 20 мкм.
Мал. б. Рибосоми:
1. 2 — відповідно велика і мала субодиниці рибосоми;
З, 4 — інформаційна (матрична) РНК;
5 — поліпептидний ланцюг;
6 — транспортна РНК з амінокислотою
(стрілкою показано напрямок руху іРНК)
Вони є в усіх клітинах, за винятком бактерій і синьо – зелених водоростей. Кількість мітохондрій у клітинах різна — від однієї (у деяких водоростей) до тисячі (печінка). Виявлено, що за певних умов (наприклад, під час голодування) число мітохондрій може зменшуватися. Спостерігаючи під мікроскопом за живими клітинами, можна побачити, що мітохондрії переміщуються в цитоплазмі, можуть утворювати скупчення. У цитоплазмі мітохондрії розмножуються шляхом поділу, можуть також утворюватись внаслідок перетворення мембран ендоплазматичної сітки.
Під електронним мікроскопом було виявлено, що мітохондрії вкриті двома мембранами (зовнішньою і внутрішньою), простір між якими заповнений рідиною. Зовнішня мембрана гладенька. Внутрішня мембрана робить впинання або складки всередину мітохондрії — кристи (лат. сrista — гребінь, виріст), які збільшують внутрішню поверхню. У більшості мітохондрій вони розміщені в поперечному напрямку, деякі з них розгалужені. Кількість крист у мітохондріях різних клітин неоднакова: їх може бути від кількох десятків до кількох сотень, причому особливо багато їх у мітохондріях клітин, які активно функціонують. Внутрішня порожнина мітохондрії заповнена напіврідкою речовиною. В ній містяться рибосоми, РНК і ДНК. На поверхні крист розміщена велика кількість ферментів, які забезпечують перебіг складних біохімічних реакцій. Функція мітохондрій полягає в тому, що в них відбуваються окисно-відновні процеси, внаслідок яких розщеплюються вуглеводи, амінокислоти та інші органічні сполуки. При цьому вивільняється енергія, яка акумулюється шляхом утворення фосфатних зв'язків в аденозинтрифосфорній кислоті — АТФ. Завдяки здатності до нагромадження АТФ мітохондрії є своєрідними акумуляторами енергії клітини. Синтезована в мітохондріях АТФ вільно виходить у цитоплазму й далі прямує до ядра та органел клітини, де в міру потреби розщеплюється і забезпечує їх енергією.
Крім АТФ у мітохондріях синтезуються власні білки, РНК і ДНК.
Пластиди — органели, специфічні для рослинних клітин, а в клітинах тваринних організмів, бактерій, синьо – зелених водоростей і грибів їх немає. У клітинах вищих рослин міститься від 10 до 200 пластид розміром 3—10 мкм, більшість із них має форму двоопуклої лінзи (трапляються у формі паличок, пластинок, лусок і зерен).
Залежно від характеру пігменту розрізняють: хлоропласти (гр. сhioros — зелений) — зеленого кольору, хромопласти — жовтого, оранжевого і червоного кольорів і лейкопласти — безбарвні пластиди. У процесі розвитку рослин пластиди одного типу можуть перетворюватися на пластиди іншого типу. Це явище поширене в природі й особливо помітне під час достигання плодів, коли змінюється їхнє забарвлення. У більшості водоростей пластиди представлені хроматофорами (у клітині він зазвичай один, значних розмірів і має форму сітки, чаші, спіральної стрічки або зірчастої пластинки).
Внутрішня будова пластид досить складна (мал. 6). У хлоропластах є власні рибосоми, ДНК, РНК, включення жиру, зерна крохмалю. Зовні хлоропласти вкриті двома мембранами, всередині заповнені напіврідкою речовиною (строма), в якій містяться особливі, властиві тільки хлоропластам структури — грани. Грани (розміром близько 1 мкм) — пакети круглих плоских мішечків (тилакоїдів), складених подібно до стовпчика монет перпендикулярно до широкої поверхні хлоропласта. Тилакоїди сусідніх гран сполучені між собою мембранними каналами (між гранні ламели) в єдину взаємозв'язану систему. Хлорофіл розміщений у певному порядку на поверхні і в товщі цих мембранних структур хлоропласта. Число гран у хлоропластах різне. Наприклад, у клітинах овочевої культури шпинату кожний хлоропласт містить 40—60 гран.
Подібно до інших органел хлоропласти не закріплені в певних місцях, а здатні змінювати своє положення в клітині шляхом пасивного переміщення разом з течією цитоплазми або шляхом активного орієнтованого переміщення. Активний рух хлоропластів особливо чітко простежується в разі значного посилення однобічного освітлення. При цьому хлоропласти скупчуються біля бічних стінок клітини й орієнтуються до джерела світла ребром. У разі слабкого освітлення хлоропласти орієнтуються більшою площиною до світла й розміщуються вздовж стінки клітини, яка обернена до світла.
За освітлення середньої сили вони займають серединне положення. Цим забезпечуються найсприятливіші умови для процесу фотосинтезу.
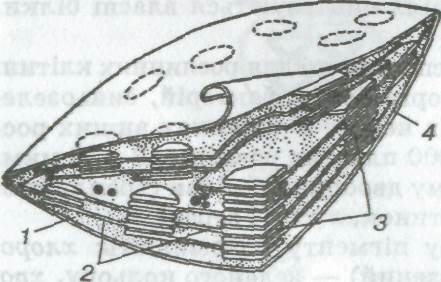
Мал. 6. Схема субмікроскопічної будови хлоропласта:
1 — строма; 2 — ламели; 3 — грани; 4 — оболонка
Складна внутрішня просторова організація структурних елементів хлоропласта сприяє ефективному поглинанню і використанню променистої енергії, а також розмежовує в часі і просторі численні й різноманітні реакції» які в сукупності становлять процес фотосинтезу. Відомо, що залежні від світла (світлові) реакції цього процесу відбуваються лише в тилакоїдах, а біохімічні (темнові) реакції — в стромі хлоропласта.
Молекула хлорофілу дуже подібна до молекули гемоглобіну й відрізняється від останньої в основному тим, що розміщений у центрі молекули гемоглобіну атом заліза замінений у молекулі хлорофілу на атом магнію.
У природі трапляються чотири типи хлорофілу: а, Ь, с, й. Хлорофіли а і Ь містяться у вищих рослинах і зелених водоростях, діатомові водорості містять хлорофіли а і с, червоні — а і d. Краще за інші вивчено хлорофіли а і Ь (вперше їх розділив російський учений М. С. Цвєт на початку XX ст.).
Крім цих хлорофілів існує чотири види бактеріохлорофілів — зелених пігментів пурпурових і зелених бактерій — а, Ь, с, d. Більшість фото синтезуючих бактерій містять бактеріохлорофіл а, деякі — бактеріохлорофіл Ь, зелені бактерії — c i d. Хлорофіл має здатність досить ефективно поглинати променисту енергію і передавати її іншим молекулам. Завдяки цій здатності хлорофіл є єдиною структурою на Землі, яка забезпечує процес фотосинтезу. Пластидам, подібно до мітохондрій, властива автономія в клітині. Вони мають свою власну спадкову інформацію, закодовану у вигляді послідовності нуклеотидів кільцевої молекули ДНК, як і у прокаріотів. Вони мають і власний білоксинтезуючий апарат, за допомогою якого синтезують специфічні білки, що входять до складу їхніх мембран. Як і мітохондрії, пластиди розмножуються поділом.
Поряд із фотосинтезом у хлоропластах відбувається синтез інших речовин, таких як білки, ліпіди, деякі вітаміни.
Завдяки наявності в пластидах ДНК вони відіграють певну роль у передаванні спадкових ознак (цитоплазматична спадковість).
Комплекс Гольджі, або внутрішній сітчастий апарат (див. мал, 1, поз. 7), під оптичним мікроскопом має форму сітки або зігнутих паличкоподібних тілець, розміщених навколо ядра. Трапляється в усіх клітинах тварин і рослин. Особливо добре його видно в нервових клітинах. Під електронним мікроскопом виявлено, що ця органела утворена гладенькими мембранами, які розміщені паралельно одна одній і утворюють систему трубочок з міхурцями різного розміру на кінцях. Розміри дрібних міхурців — 20—30 нм, великих — до 2000 нм.
Основна функція комплексу Гольджі — виведення синтезованих клітиною речовин. Ці речовини транспортуються по канальцях ендоплазматичної сітки і нагромаджуються у міхурцях комплексу Гольджі. Звідси вони або виводяться з клітини назовні, або використовуються в процесі життєдіяльності клітин. У комплексі також відбувається концентрування речовин (наприклад, барвників), які надходять у клітину ззовні і мають бути виведені з неї. Крім того, комплекс Гольджі бере участь у синтезі тих хімічних сполук, з яких будується клітинна мембрана.
Лізосоми (див. мал. 1, поз. 6; мал. 4, поз. 4, 5; гр. lysis — розчинення) — дрібні, видимі тільки під електронним мікроскопом органели діаметром близько 1 мкм. Вони вкриті щільною мембраною і містять до 40 різних ферментів, здатних розщеплювати білки, жири і вуглеводи. Кількість лізосом у клітинах різна. Особливо багато їх (до кількох сотень) у клітинах, здатних до фагоцитозу. Функція лізосом полягає у перетравленні речовин, які потрапили в клітину у процесі фаго- або піноцитозу, а також у руйнуванні окремих органел або клітини у разі ЇЇ відмирання. Це відбувається внаслідок руйнування оболонки лізосом і вивільнення з неї ферментів. Іноді ферменти лізосом беруть участь у руйнуванні міжклітинної речовини, а також цілих органів. Наприклад, під дією ферментів лізосом відбувається розсмоктування хвоста у пуголовків жаби в процесі метаморфозу. Утворення нових лізосом пов'язане з міхурцями комплексу Гольджі.
|
|
|
|
|
Дата добавления: 2015-05-23; Просмотров: 4407; Нарушение авторских прав?; Мы поможем в написании вашей работы!