КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Зміна алеля дикого типу та його продуктів
|
|
|
|
(mPHK і поліпептидного ланцюга) в результаті вставки та делеції
| Структури | Частина послідовності | |
| Алель дикого типу | ДНК m PHK | -C-C-C-G-G-T-A-G-C-C-C-C- -G-G-G-C-C-A-T-C-G-G-G-G- -C-C-C-G-G-U-A-G-C-C-C-C- |
| Поліпептидний ланцюг | — Pro — Gly —Ser —Prо — | |
| Після вставки | ДНК | +С -C-C-C-C-G-G-T-A-G-C-C-C-C- -G-G-G-G-C-C-A-T-C-G-G-G-G- -C-C-C-C-G-G-U-A-G-C-C-C-C- |
| m PHK | ||
| Поліпептидний ланцюг | — Pro — Arg — amber (кінець ланцюга) | |
| Після делеції | ДНК | -G -C-C-C-G-T-A-G-C-C-C-C- -G-G-G-C-A-T-C-G-G-G-G- -C-C-C-G-U-A-G-C-C-C-C- |
| m PHK | ||
| Поліпептидний ланцюг | — Pro —Val — Ala —Pro |
Більшість генних мутацій дають негативний ефект й обумовлюють випадіння якої-небудь ферментативної активності шляхом спадкоємного захворювання, можуть сильно розрізнятися по виразності симптомів хвороби. У диплоїдів мутації виникають тільки в одному із двох алелів. У результаті виходять гетерозиготи, у яких фенотип визначається взаємодією алелей.
Індукувати генні мутації можуть ультрафіолетові промені; іонізуюче випромінювання, хімічні мутагени. З хімічних мутагенів найчастіше застосовують алкілуючі агенти. До них відносяться: етилметансульфонат, нітрозогуанідинові сполуки, аналоги азотистих основ, такі як бромурацил та 2-амінопурин, а також азотиста кислота та гідроксиламін. Мутації зі "зрушенням рамки" індукуються акридином.
Тільки в деяких випадках мутації реалізуються відразу ж після впливу фізичного або хімічного мутагену. Частина ушкоджень усувається в результаті репаративних процесів. При безпомилковій репарації знову відновлюється нормальний вихідний стан, тоді як помилка при репарації може, наприклад, привести до заміни основи. Можна припустити наступні шляхи виникнення генних мутацій:
а) одна основа перетворюється в іншу ( замінаоснов);
б) ДНК змінюється так, що починається процес її репарації, при якому включається «не та» основа;
в) невідповідний нуклеотид включається в результаті помилки реплікації.
Мутації виникають із частотою 10-5-10-10 (на один ген). З такою же частотою деякі з мутантів можуть знову перетворюватися в дикий тип. Або нова зміна усувається в результаті справжньої зворотної мутації, або ознака дикого типу відновлюється внаслідок другої, так називаної супрессорної, мутації.
Виникнення мутацій при реплікації залежить від частоти помилок при включенні комплементарних нуклеотидов ДНК-полімеразою. Такі мутації зустрічаються дуже рідко.
В інтактних клітинах існують різні «ремонтні» системи, що усувають ушкодження, викликані опроміненням або хімічними мутагенами. У репаративних процесах розрізняють фотореактивацію, ексцизійну репарацію й постреплікативну репарацію.
Найкраще вивчена репарація ушкоджень, викликаних ультрафіолетовими променями (репарація інших ушкоджень частково відбувається аналогічним чином). При опроміненні ультрафіолетом між сусідніми піримідиновими основами одного ланцюга виникають димери, частіш усього димер Т-Т, тобто замість водневих зв'язків між Т та А двохнуклеотидних ланцюгів утворюються зв'язки Т-Т усередині одного ланцюга (рис. 10.4). Фотореактивація відбувається у разі впливу видимого світла. При цьому репаративний фермент розділяє димер на мономери й знову відновлює водневі зв'язки Т-А між комплементарними ланцюгами.
Ексцизійна й постреплікативна репарація не залежить від світла, і тому її називають також темновою репарацією.
При ексцизійній репарації (лат. excisio – вирізання; рис. 10.4) вирізується ушкоджена ділянка ДНК. Спочатку едонуклеаза розрізає один ланцюг. Наступний фермент – екзонуклеаза – видаляє змінену частину, а ДНК-полімераза знову синтезує в напрямку 5'®3' відсутню ділянку, комплементарний до антипаралельного ланцюга ДНК. І, нарешті, вільні кінці старої частини ланцюга з'єднуються з кінцями знову синтезованої ділянки за допомогою лігази. Фотореактивація та описана тут ексцизійна репарація коротких ділянок ДНК протікають без помилок і, отже, не ведуть до мутацій.
Якщо димери не будуть усунуті, то відповідні основи не зможуть виконувати роль матриці, і в цих місцях у знову синтезованому ланцюзі ДНК виявляться пропуски. Шляхом рекомбінації між двома подвійними ланцюгами ДНК – продуктами реплікації – можливе утворення одного нормального подвійного ланцюга (постреплікативна репарація). Коли після реплікації в результаті такої рекомбінації виходить нова інтактна подвійна спіраль ДНК, процес репарації теж не веде до появи помилок. Якщо ушкодження лежать так тісно одне біля одного, що пропуски перекриваються, тоді для заповнення пропусків використовується інша «ремонтна» система (SOS- репарація), здатна синтезувати новий ланцюг ДНК і на дефектній матриці. При цьому нові основи включаються таким чином, що можуть виникнути мутації (рис. 10.4).
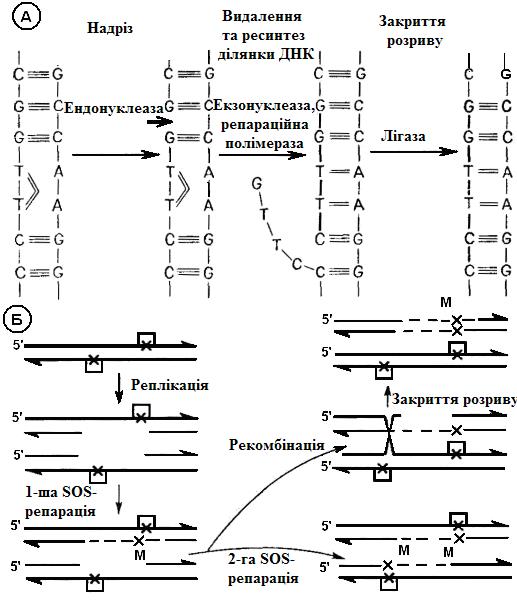
Рис 10.4. Репаративні процеси
А.Ексцизійна репарація (на прикладі Escherichia coli).
Б. Постреплікативна репарація. У наданому прикладі розрив в одній молекулі ДНК закривається шляхом SOS- репарації, причому виникає мутація (М). У другій молекулі ДНК розрив може бути теж заповнений шляхом SOS- репарації або закритий шляхом рекомбінації з наступним репаративним синтезом, при якому матрицею служить інтактний ланцюг ДНК. (За Bohme, Adler, зі змінами)
Більшість мутантів поступаються дикому типу в пристосованості, менш життєздатні й тому відсіваються в процесі добору. Для селекції й еволюції велике значення мають порівняно рідкі мутанти зі сприятливими або нейтральними змінами. Щоб мати достатнє число мутантів, можна за допомогою мутагенів підвищувати частоту мутацій до 10-2.
Людина, навпроти, повинна оберігати себе від несприятливих змін, до яких можуть приводити мутації. Кожна мутація, що зачіпає тканину, з якої утворюються статеві клітини, буде передаватися наступним поколінням.
10.2 Рекомбінації
Під рекомбінацією розуміють обмін алелями, перерозподіл структур, що несуть генетичну інформацію, з'єднання їх у нові сполучення. Передумовою для здійснення рекомбінацій є об'єднання різного генетичного матеріалу. Розрізняють два основних типи такого об'єднання:
1) статевий процес, при якому відбувається злиття генеративних клітин й їхніх ядер;
2) парасексуальний процес, коли відбувається злиття вегетативних клітин, рекомбінація у вегетативних клітинах або перенос частини геному.
Перенос частини геному відбувається у разі:
а) кон'югації у бактерій – контакті між клітинами, при якому відбувається передача плазмід або частин геному, активованих плазмідами;
б) трансдукції – переносі генетичного матеріалу клітини вірусами;
в) трансформації – передачі ДНК через позаклітинне середовище;
г) рекомбінації між вірусними геномами.
Об'єднання різного генетичного матеріалу в основному можливо лише в межах одного виду або близько родинних видів. Наявність бар'єрів для схрещування обумовлює збереження видів.
В еукаріот після об'єднання різного генетичного матеріалу в одному ядрі утворюються гетерозиготні клітини, щонайменше із двома геномами, що розрізняються за алелями одного або декількох генів. Такі організми називають помісями або гібридами. У прокаріот після переносу генів клітини виявляються диплоїдними тільки по одному гену або якійсь частині геному і у випадку розходження в алелях називаються гетерогенотами.
Гібридний стан може зберігатися протягом ряду поколінь, або ж відбувається рекомбінація. Рекомбінація може бути:
а) на рівні цілих хромосом – з утворенням нового сполучення між ними;
б) усередині хромосом – між їхніми частинами;
в) між геномом і плазмоном або плазмідами, вірусами й іншими додатковими послідовностями ДНК.
10.2.1 Рекомбінація цілих хромосом
Цей процес визначається двома фазами:
1) злиттям гаплоїдних (у нормі) клітин у диплоїдну зиготу;
2) координованим розподілом хромосом у мейозі.
Ці дві ступені визначають характерні типи розщеплення після схрещувань. Уперше ці типи спостерігав Мендель ще у 1865 р., і після їхнього повторного відкриття Коренсом, Чермаком: і де Фризом (у 1900 р.) вони були покладені в основу класичної генетики.
У разі монофакторіального спадкування вихідні індивідууми; (батьки – Р) розрізняються алелями одного гена. Різні алелі одного гена завжди позначають однаковим головним-символом (наприклад, а), щоб відзначити їхню приналежність до одного генного локусу. Нормальний алель, тобто алель дикого типу, позначають знаком «плюс» (а+) або А, а мутантний алель знаком «-» (а–) або а.
Прикладом монофакторіального спадкування з гомозиготними батьками є спадкування фенілкетонурії (рис. 10.5). Нормальний алель позначимо а+,а мутантний алель, що обумовлює дефект ферменту й тим самим хворобу, — а–. У нашому прикладі батько гомозиготний з двома нормальними алелями, тобто а+а+, а мати гомозиготна з двома аномальними алелями, тобто а–а–.
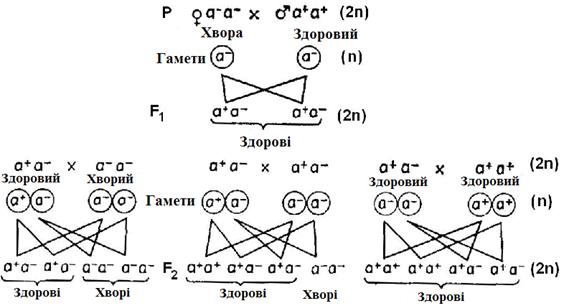
Рис. 10.5. Монофакторіальне спадкування.Приклад: рецесивна спадкоємна хвороба. а + – нормальний алель; а– – мутантний алель,
що обумовлює хворобу
Вона здорова тільки фенотипово завдяки правильній дієті. Таким чином, батьки розрізняються за алелями одного гена (інші розходження ми заради простоти не враховуємо). Це відноситься до всіх соматичних клітин батьків. При утворенні статевих клітин диплоїдне число хромосом 2 n =46 зменшується до гаплоїдного n =23. Алелі разом з гомологічними хромосомами розподіляються так, що всі батьківські гамети містять алель а+, усі материнські – алель a -. При заплідненні ядра гамет зливаються й утворюють диплоїдну зиготу. Алелі а+ й а– об'єднані тепер в одному ядрі, що стає гетерозиготним (а+а–). З такої зиготи у результаті мітозів розвивається гетерозиготний ембріон і народжується дитина F1 (перше покоління потомства). Фенотипово дитина здорова, тому що нормальний алель а+ домінує над рецесивниммутантним алелем а-. Зважаючи на те, що всі комбінації гамет відносно даного гена повинні бути однакові, то всі діти від цього шлюбу гетерозиготні і здорові, хоча мати гомозиготна й хвора.
Нащадки представників фенотипово здорового, але гетерозиготного покоління F1 залежать від вибору шлюбного партнера. При гетерозиготних батьках (а+а– а+а–) спадкоємність буде такою ж, як при схрещуванні гібридівF1. У особин F1 всі соматичні клітини мають генотип а + а–. У мейозі з однаковою частотою утворюються гамети а+ й а–. При утворенні зигот гамети можуть із однаковою імовірністю зливатися в наступних сполученнях (першою зазначена чоловіча гамета, другою – жіноча): 1) а+ з а+; 2) а+ з а–; 3) а– з а+; 4) а– з а–. Тому потомство F2 розщеплюється у відношенні 1 а+а+: 2 а+а–: 1 а-а–.
Схрещування між гетерозиготами (а+а–) і відповідними гомозиготами (а+а+ або а-а–) називають поворотними схрещуваннями.
Таким чином, як вище було пояснено на прикладі фенілкетонурії, у диплоїдних рослин і тварин, включаючи людину, успадковуються всі ознаки, обумовлені одним геном (монофакторіальне) з домінантним і рецесивним алелями. При проміжному спадкуванні відбувається аналогічне розщеплення за генотипом, але фенотипи гетерозигот займають проміжне положення між фенотипами батьків.
Мендель, ґрунтуючись на результатах своїх експериментів по схрещуванню, сформулював закономірності, відомі наразі як «закони Менделя».
Перший закон Менделя(закон однаковості F1): при схрещуванні гомозиготних батьківських форм у першому поколінні потомства всі особини однотипні (рис. 10.5).
Другий закон Менделя(закон розщеплення): після схрещування нащадків F1 двох гомозиготних батьків у поколінні F2 відбувається закономірне розщеплення (рис. 10.5). При розходженні по алелям одногогена F2 розщеплюється за генотипом у відношенні 1:2:1. Розщеплення за фенотипом залежить від взаємодії алелей. У більшості випадків ознака, обумовлена алелем дикого типу, домінує над мутантними алелями, рідше навпаки. Крім того, існують усі переходи від домінантності через неповне домінування до проміжного співвідношення, при якому обидва алеля проявляються однаковою мірою і фенотип виявляється «усередненим». При монофакторіальному спадкуванні потомство розщеплюється за фенотипом у випадку повного домінування у відношенні 3:1, а при неповному домінуванні – у відношенні 1:2:1.
Третій закон Менделя (закон незалежного розподілу): алелі кожного гена розподіляються у потомстві незалежно від алелей інших генів.
Цей закон справедливий тільки для генів, що перебувають або в різних хромосомах, або в одній хромосомі, але досить далеко один від одного (50 або більше одиниць карти). Точного розщеплення за Менделем можна чекати тільки тоді, коли аналізоване потомство досить велике.
У статевих хромосомах локалізуються не тільки гени, необхідні для розвитку первинних і вторинних статевих ознак. У Х хромосомах перебувають, наприклад, гени кольорового зору (дефект: червоно-зелена сліпота, або дальтонізм) і згортання крові (дефект: гемофілія), що передаються у спадщину разом з Х -хромосомою. У Y-хромосомі відповідних алелей немає, і тому рецесивні гени, що перебувають у Х -хромосомі, у особин чоловічої статі проявляються у фенотипі (гемізиготність).
10.2.2 Внутрішньохромосомна рекомбінація
При внутрішньохромосомній рекомбінації в галоїдних і диплоїдних еукаріот гомологічні хромосоми взаємно обмінюються ділянками хроматид. У бактерій і вірусів частини геному замінюються гомологічними (алельними) ділянками.
Якби рекомбінація була обмежена лише появою нових сполучень хромосом, то всі гени, що перебувають в одній хромосомі, успадковувалися б разом. Гени, що знаходяться у одній хромосомі називають зчепленими. При повному зчепленні двох генів спостерігалося б розщеплення, представлене на рис. 10.6, A. Таке розщеплення зустрічається тільки у випадку тісно зчеплених генів, тому що в профазі мейозу у процесі кросинговеру відбувається взаємний обмін частинами хроматид. Аналогічний обмін можливий і у мітозі – так званий мітотичний кросинговер. У результаті кросинговера в потомстві з'являються рекомбінанти (рис. 10.6, Б).
Чим більше віддалені один від одного два гени в одній хромосомі, тим більше імовірність того, що між ними відбудеться кросинговер. Частота рекомбінацій зростає зі збільшенням відстані між генами, і тому її використовують як міру цієї відстані: за «одиницю карти», або морганіду приймають відстань, що відповідає одному відсотку рекомбінації. Гени, віддалені один від одного більше чим на 50 одиниць карти, поводять себе як не зчеплені. Рекомбінації можуть відбуватись і проміж різними мутантними ділянками у межах одного гена, але це буває значно рідше.
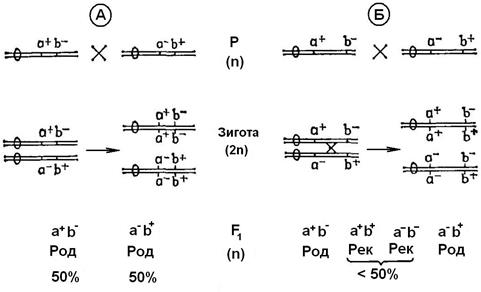
Рис. 10.6. Спадкування зчеплених генів у гаплоїдів: А– без кросинговера
Б – із кросинговером. Подвійна лінія з овалом – хромосома з двох хроматид із центромерою. Род – батьківський – тип; Рек –рекомбінантний тип
Бактеріальна клітина може стати донором генетичного матеріалу, якщо вона містить кон’югаційну плазміду. Перенос бактеріальних генів фагами називається трансдукцією. Трансформацією називають процес, при якому позаклітинна ДНК проникає у клітину реципієнта і вбудовується в геном.
Запитання для самоперевірки
1. Види спадкоємних змін.
2. Типи мутацій.
3. Мутації типа зміни плоїдності та їхні наслідки.
4. Хромосомні мутації.
5. Генні мутації.
6. Мутагени та типи, викликаних ними ушкоджень.
7. Репаративні процеси.
8. Що розуміють під рекомбінацією?
9. Чим визначається процес рекомбінації цілих хромосом?
10. Монофакторіальне спадкування. Фенотип та генотип лершого покоління потомства гомозиготних батьків, що розрізняються алелями одного гена.
11. Монофакторіальне спадкування. Фенотип та генотип другого покоління потомства гомозиготних батьків, що розрізняються алелями одного гена.
12. Закони Менделя.
13. Внутрішньохромосомна рекомбінація.
14. Рекомбінація у бактерій трансдукцією.
15. Трансформація та трансдукція.
Лекція 16
ГЛАВА 11
|
|
|
|
|
Дата добавления: 2015-05-24; Просмотров: 854; Нарушение авторских прав?; Мы поможем в написании вашей работы!