КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Збудливість – рух – поведінка
|
|
|
|
План
9.1 Збудливість (подразливість)
9.2 Рух (рухливість)
9.3 Поведінка
9.1 Збудливість (подразливість)
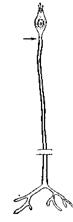
Невід'ємною властивістю всіх живих систем є збудливість – здатність реагувати на певні впливи зовнішнього середовища (подразники) тимчасовими змінами, найхарактернішими з яких є зміні електричного потенціалу між внутрішньою та зовнішньою сторонами клітинної мембрани. У тварин є певні клітини, які спеціалізовані для сприйняття подразників – рецептори (рис. 9.1).
В місці входу подразнення (яке впливає ззовні) веде до збудження, що пов'язане зі зміною мембранного потенціалу. Це не просте перетворення енергії подразника у збудження. Подразник (стимул) служить лише пусковим або керуючим фактором.
9.1.1 Потенціал спокою
У всіх легко збудливих клітинах існує помітний електричний потенціал між внутрішньою стороною плазматичної мембрани й поверхнею клітини. Цей мембранний потенціал спокою має завжди одну спрямованість (внутрішня сторона негативна стосовно зовнішньої) і приблизно однакову величину: від -50 до -90 мВ (у рослин до -200 мВ). Він є основою збудливості клітини.
| 
|
| А | Б |
Рис. 9.1. Сенсорні нейрони (схема).
А. Нюхальна клітина (хребетного). Б. Дотикальна клітина шкіри (хребетного). Стрілкою показане місце виникнення потенціалу дії
Мембранний потенціал спокою обумовлений – 1) асиметричним розподілом іонів між внутрішнім середовищем клітини й позаклітинною рідиною і 2) специфічними властивостями проникності плазматичної мембрани. Концентрація К+ усередині клітини значно вище, ніж зовні, а іонів Na+, навпаки, зовні більше, ніж усередині. З аніонів поза тваринною клітиною найбільше С1–, а усередині клітини – SО42– і білкових аніонів. Плазматична мембрана тваринної клітини у «спокої» добре пропускає К+, гірше – С1– і зовсім погано – Na+ (у випадку гігантського аксона кальмара відносні константи проникності для К+, С1– і Na+ рівні відповідно 1, 0,45 та 0,04). У першому наближенні потенціал спокою тваринної клітини відповідає потенціалу рівноваги для К+. Іони К+ через існуючий градієнт концентрації (хімічнй градієнт) прагнуть виходити назовні, але цьому протидіють усередині аніонні партнери, що залишаються. У результаті створюється електричний градієнт (зовні – надлишок, усередині клітини – недолік позитивних зарядів.
9.1.2 Збудження
Зменшення потенціалу спокою під час деякого впливу називають деполяризацією. Величина деполяризації залежить від інтенсивності подразнення. Після припинення стимуляції деполярізація більш-менш швидко падає та залишається обмеженою місцем свого виникнення (локальна відповідь). Якщо деполяризація перевищує певний критичний рівень (критичний мембранний потенціал, мембранний поріг), починаються процеси, що ведуть до появи потенціалу дії (спайка, або пік-потенціалу).
Потенціал дії складається з фази, звичайно дуже швидкої, повної деполяризації мембрани з наступною фазою інверсії полярності (зовнішня сторона ненадовго стає електронегативною стосовно внутрішньої). Після цього відбувається відновлення нормальної поляризації – нормального потенціалу спокою. Протікання і швидкість цієї реполяризації в різних клітин різні. На своїй вершині потенціал дії досягає рівня, за знаком і величиною близького до рівноважного потенціалу для Na+.
Це пояснюється тим, що подразнення веде до сильного підвищення проникності для Na+ (Na+-активація). Вона стає приблизно в 20 разів вище, ніж для іонів К+. У результаті іони Na+ пасивно переходять у клітину. Коли цей процес припиняється, а також на якийсь час зростає проникність для К+, наступає, нарешті, фаза спаду потенціалу дії. Під час деполяризації та інверсії полярності, а також на початку реполяризації відповідна ділянка мембрани «рефрактерна», тобто тимчасово не здатна до збудження. Цей період називається періодом абсолютної рефрактерності. Після цього збудливість поступово відновлюється.
На відміну від локальної відповіді потенціал дії або виникає на повну силу (з максимальною амплітудою), або не виникає зовсім: він підкоряється закону «все або нічого». Так само як й амплітуда, тривалість потенціалу дії завжди однакова незалежно від сили подразнення.
9.1.3 Проведення збудження
Проведення потенціалу дії (проведення збудження) пов'язане із процесами, що відбуваються в місці дуже тонкої мембрани нервового волокна. У мімози та у деяких інших рослин, які виявляють подразливість, потенціали дії передаються від клітини до клітини головним чином через витягнуті паренхімні клітини флоеми та протоксилеми зі швидкістю 2-5 см/с. У нервовій системі людини досягаються швидкості в 100-150 м/с.
У немієлінізованих волокнах, відповідно до теорії місцевих токів, збудження проводиться у такий спосіб. Зміна знака мембранного потенціалу у збудженій ділянці (зовнішня сторона стає електронегативною стосовно внутрішньої) спричиняє появу токів, що вирівнюють потенціал із сусідніми, ще не збудженими ділянками (рис. 9.2). Ці ділянки настільки деполяризуються, що і тут досягається критичний граничний потенціал і у результаті виникає потенціал дії, а в зоні, збудженій спочатку, тим часом, відновлюється потенціал спокою. Завдяки повторенню цього процесу на усіх нових ділянках, збудження безупинно поширюється уздовж нервового волокна. Оскільки потенціал дії створюється на кожній ділянці мембрани заново, хвиля збудження йде волокном, не слабшуючи, без загасання. Повернути назад вона не може, тому що за кожним потенціалом дії знаходиться зона, що перебуває в рефрактерному стані.
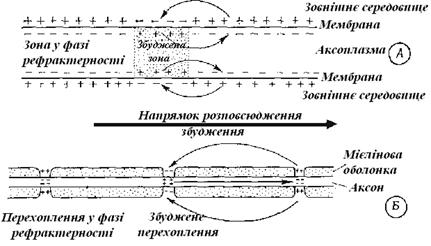
Рис. 9.2.Проведення збудження
А. Безперервне у безмієлінових волокнах. Б. Сальтаторне у мієлінових волокнах
У мієлінізованих волокнах на ділянках мембрани, покритих ефективним ізолятором – мієліновою оболонкою, процес збудження відбуватися не може. У цьому випадку потенціали дії можуть виникати тільки там, де оболонка переривається – у перехопленнях Ранв’є, розташованих на певних відстанях один від одного (рис. 7.8). Таким чином, збудження перескакує від одного перехоплення до іншого – це, так називане, сальтаторне (стрибкоподібне) проведення збудження. Перевагами такого механізму є: а) більша швидкість проведення, б) економія метаболічної енергії та в) підвищена надійність, тому що щільність токів на перехопленнях досягає більшої величини.
9.1.4 Синаптична передача збудження. З'єднання нейронів
Збудження може передаватися з одного нейрона на іншій у синапсах. При цьому йому доводиться переходити синаптичну щілину, що має в різних синапсах різну ширину. Більшість синапсів передає збудження тільки в одному напрямку (вентильна функція синапсу) – від пресинаптичної клітини до постсинаптичної. Один нейрон може мати сотні синапсів.
У разі електричної передачі збудження, що є вже у рослинних клітинах (але без синапсів), наприклад у мімози, кожен потенціал дії, приходить до синапсу, безпосередньо, як подразник, збуджує постсинаптичну клітину, викликаючи в ній потенціал дії.
У разі більш розповсюдженої хімічної передачі збудження потенціал дії, що прийшов до синапсу, викликає вивільнення із пресинаптичої клітини у синаптичну щілину специфічної речовини (медіатора). Ця речовина дифундує до лежачої навпроти мембрани постсинаптичної клітини, де зв'язується зі специфічними молекулами-рецепторами. Комплекс, що утворюється, медіатор – рецептор викликає зміни проникності субсинаптичної мембрани, і тим самим – зрушення мембранного потенціалу. Ця «локальна відповідь» може виражатися у деполяризації, що у разі достатньої величини викликає потенціали дії (збуджуючий постсинаптичний потенціал – ЗПСП), або ж у гіперполяризації, що утруднює або виключає виникнення потенціалів дії (гальмівний постсинаптичний потенціал – ГПСП). Після цього медіатор, що виділився, негайно ж вилучається за допомогою спеціальних механізмів.
Найбільш відомий медіатор – ацетилхолін. Він забезпечує, наприклад, передачу збудження з нервів на поперечносмугасті м'яза у хребетних. У синаптичній щілині ацетилхолін швидко інактивується ацетилхолінестеразою, що розщеплює його на холін та ацетат. Речовинами, що придушують активність ацетилхолінестерази (наприклад, фізостігміном), можна блокувати нервово-м'язову передачу збудження. Кураре діє по-іншому – ця отрута робить постсинаптичну мембрану нечуттєвою до ацетилхоліну, займаючи місце ацетилхоліну
на рецепторах. Інші відомі медіатори – адреналін, норадреналін, аміномасляна кислота, серотонін, глутамат, гліцин, дофамін та ін.
Однією з важливих форм з'єднання нейронів у нервовій системі є рефлекторна дуга. Вона складається з рецептора з його аферентним (сенсорним) провідним шляхом та еферентного провідного шляху з ефектором (рис. 9.3).
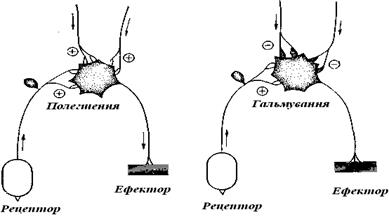
Рис. 9.3. Лабільність рефлекторних дуг.Полегшення та гальмування рефлексу в синапсі під впливом двох нейронів, що не входять у дану дугу
У найпростішому випадку збудження з аферентного нейрона прямо передається на еферентний (моносинаптичні рефлекси); але в більшості випадків передача йде через вставні нейрони (полісинаптичні рефлекси). Рефлекс – це реакція, яка здійснюється нервовим механізмом, на специфічний подразник, що регулярно повторюється більш-менш однаковим чином. Рефлекторні дії залежно від тих або інших умов можуть виконуватися по-різному (пластичність). Наприклад, якщо ми перешкодимо спинальній (позбавленої головного мозку) жабі потерти спину однією ногою, то до роздратованого місця на шкірі потягнеться інша нога.
9.1.5 Навчання та пам'ять
Про навчання ми говоримо, коли в певній ситуації, що сприймається органами почуттів, імовірність певної форми поводінки змінюється в результаті попередніх зустрічей з тією ж ситуацією. Деякі автори додають: змінюється накраще, «а саме так, що організм виявляється краще пристосованим до свого середовища». Навчання найтіснішим чином пов'язане з пам'яттю: у процесі навчання відбувається перенос інформації із зовнішнього середовища в сховище пам'яті. Здатність до навчання з безсумнівністю встановлена вже у нижчих черв’яків (планарій).
Розрізняють наступні типи процесів навчання:
1. Звикання (габітуація). Тварини, що повторно піддаються впливу того самого подразника, який не визиває ніяких біологічно значимих подій, реагують на нього усе слабкіше та, зрештою, зовсім перестають реагувати. Так, через звикання припиняється «сторожкість» кішки або «завмирання» пташеняти під час певних звукових сигналів, спурхування шпаків з вишневого дерева при пострілі та ін. Звикання не можна пояснити стомленням, воно скоріше пов'язане з нагромадженням гальмування, що зупиняє реакцію.
2. Імпринтинг (запам'ятовування). Це процес навчання, часто необоротний, що протікає надзвичайно швидко, протягом більш-менш короткого «критичного періоду». Наприклад, реакція слідування у виводкових птахів необоротно закріплюється у перші дні життя. Фаза слідування в гусячих птахів закінчується вже через 12-24 год після вилуплення. У птахів і ссавців відомо також явище статевого імпринтинга.
3. Вироблення класичних умовних рефлексів. Воно полягає у зв'язуванні «умовного» подразника (наприклад, слухового або зорового) з яким-небудь рефлексом, раніше з ним не зв'язаним (наприклад, слинним рефлексом у собаки). Якщо безумовний подразник (наприклад, їжу) пред'являють одночасно з умовним і такий дослід повторюють багаторазово, то утворюється зв'язок між обома стимулами і, зрештою, тварина починає відповідати специфічною реакцією (наприклад, виділенням слини) вже на один лише умовний подразник, без пред'явлення безумовного.
4. Вироблення інструментальної, або оперантної умовної реакції. Цю форму навчання називають також «закріпленням дій, що ведуть до успіху» або «методом проб і помилок». Тварина може вільно пересуватися в експериментальних апаратах (так званих ящиках Скиннера, лабіринтах, пристроях для дослідів з вибором об'єктів тощо) і при цьому набирається досвіду, засвоюючи, що певні спонтанні дії винагороджуються або дозволяють уникнути неприємного «караючого» подразника.
5. Навчання шляхом наслідування. Це порівняно рідкісний тип, що зустрічається тільки у вищих хребетних.
6. Навчання у результаті «збагнення» (інсайту). Рішення поставленого завдання за допомогою нової комбінації дій приходить тут раптово. Цьому передує фаза відносної нерухомості, під час якої різні можливі дії оцінюються і порівнюються «у розумі», без випробування їх методом проб і помилок. Така форма навчання вірогідно встановлена тільки у вищих мавп та у людини.
На підставі різних спостережень – головним чином на хребетних – можна виділити, щонайменше, дві форми пам'яті, що розрізняються своїми механізмами та властивостями. Так називана короткочасна пам'ятьу людини має максимальну ємність близько 160 байт. Тривалість збереження її слідів – від декількох секунд до декількох хвилин. Інформація, що надійшла до короткочасної пам'яті, доступна відразу ж після запам'ятовування. Для цієї форми пам'яті характерна чутливість до перешкод. Інформація в ній може бути стерта в результаті переохолодження мозку, електрошоку, недостатнього постачання мозку киснем, наркозу або введення речовин, що викликають судоми (коразол та ін.). Оскільки у всіх цих випадках електрична активність мозку сильно порушується або навіть зовсім припиняється, можна припустити, що короткочасна пам'ять заснована на чисто функціональних, електрофізіологічних процесах. Можливо, що виникле збудження якийсь час циркулює замкнутими нейронними шляхами, а потім безвісти зникає, якщо інформація не переноситься в довгострокове сховище.
Довгострокова пам'ять має дуже більшу ємність – від 1010 до 1014 байт. Накопичена в ній інформація зберігається довго (іноді до кінця життя) і дуже стійка проти спроб її стерти: охолодження мозку аж до зникнення всякої вимірюваної активності не приводить до істотної втрати слідів пам'яті. Є думка, що інформація в довгостроковій пам'яті фіксується хімічним способом. Вірогідно показано, що під час навчання в певних областях мозку збільшується синтез РНК і, що із процесами навчання зв'язані зміни в синтезі білка. Загальне придушення процесів синтезу в нейронах приводить до різко виражених функціональних порушень.
Багато авторів виділяють ще дві форми пам'яті – так називану проміжну пам'ять, у якій сліди зберігаються (у пацюків) близько 4 годин, і попередню короткочасну сенсорну пам'ять із ще коротшим часом збереження слідів (кілька сотень мілісекунд).
9.2 Рух (рухливість)
Рухливість також відноситься до основних властивостей живого. Рух може відбуватися усередині живого організму і служити для транспорту речовин. Поряд із цим можливо переміщення всього тіла або його частин.
9.2.1 Ростові рухи
Ростові рухи властиві в основному рослинам і виражається в тім, що через нерівномірний ріст протилежних сторін циліндричного органа (стебла, кореня), цей орган викривлюється. Прикладом служить фототропізм – спрямована реакція скривлення, що викликається однобічним освітленням; пагони рослин, як правило, викривлюються убік світла.
Однобічне освітлення зміщає в затінену сторону потік ростового гормону ауксину, звичайно спрямованого строго униз (рис. 9.4). Збідніння ауксином освітленої сторони пагона приводить до гальмування росту, а збагачення ауксином затіненої сторони – до стимуляції росту, що й веде до скривлення.
А Б
Рис. 9.4. Фототропізм(світлі стрілки – напрямок потоку світла).
А. Позитивний фототропізм осі пагона, негативний фототропізм зародкового кореня, поперечний фототропізм листів проростка гірчиці. Б. Поперечний зсув ауксину (чорні стрілки), і скривлення колеоптиля вівсу, що починається слідом за ним
9.2.2 Тургорний рух
Тургорний рух – це оборотне скривлення, що виникає у рослин через нерівномірну зміну тургорного тиску на протилежних сторонах циліндричного органу. Приклади: денне підведення і нічне опускання листів, наприклад у бобових; дуже швидка (триваюча 0,02-1 с) реакція на механічне подразнення (сейсмонастія) у мімози, деяких комахоїдних та інших рослин.
9.2.3 Амебоїдний рух
Амебоїдний рух властивий амебам та іншим одноклітинним організмам, деяким яйцеклітинам і клітинам багатоклітинних тварин (амебоцитам, фагоцитам, лейкоцитам та ін.), а також міксоміцетам. У клітини утворюються та знову втягуються протоплазматичні відростки (псевдоподії). Амебоїдний рух служить для прийому їжі та для пересування клітин (локомоції).
9.2.4 Рух за допомогою джгутиків і війок
Джгутики і війки роблять періодичні або безладні рухи, які потрібні або для локомоції, або для того, щоб створювати плин рідини. Рух обумовлений взаємним ковзанням фібріл (результат взаємодії білків тубуліну і динеїну), які знаходяться усередині джгутика.
Під час удару окремої війки відбувається активне взаємне ковзання фібрил по всій їхній довжині одночасно, а під час зворотного руху – зворотне ковзання на обмеженій ділянці, що зміщується від початку до кінчика війки. Рух безлічі війок однієї або багатьох клітин координовані між собою.
9.2.5 М'язовий рух
М'язи складаються з м'язових волокон.Ці волокна можуть бути одноядерними або, у випадку злиття великої кількості м'язових клітин, багатоядерними. М'язовий рух заснований на здатності міофібріл, що лежать у цитоплазмі м'язового волокна, скорочуватися у поздовжньому напрямку за рахунок енергії АТР і знову подовжуватися.
У так називаній поперечносмугастій мускулатурі, розповсюдженій майже у всьому тваринному світі (кістякова мускулатура, м'язи серця і язика), у багатоядерних м'язових волокнах в світловому мікроскопі можна бачити поперемінно світлі й темні поперечні смуги. Світлі диски I (ізотропні), які мають слабку подвійну променезаломлюваність, розділені навпіл поперечною проміжною лінією – пластинкою Z (рис. 9.5).
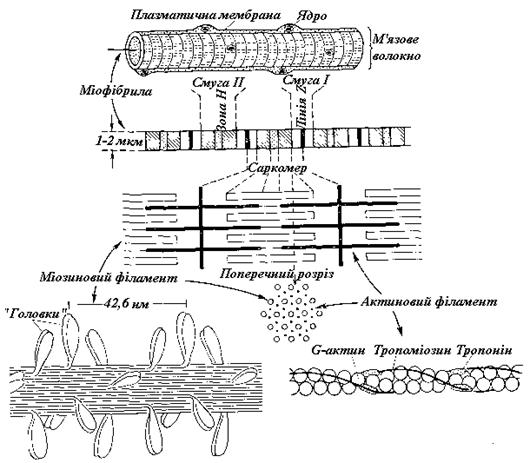
Рис. 9.5. Будова поперечносмугастого м'язового волокна
Темні диски А (анізотропні) з сильною подвійною променезаломлюваністю мають в середині трохи більше світлу зону Хензена (зону Н), що сама може бути розділена навпіл серединною лінією (лінією М). Відрізок м'язового волокна, обмежений двома пластинками Z (у м'язах теплокровних його довжина близько 2 мкм), називається саркомером.
Як показує електронна мікроскопія, міофібрили складаються з ланцюгів розташованих паралельно елементарних ниток (міофіламентів) двох типів. Більш товсті філаменти з L -міозину (діаметр 10 нм) виявляються тільки в дисках А. Навколо них групується по 6 більш тонких актинових філаментів (діаметр 6 нм); вони йдуть від самих пластинок Z і тільки усередині зон Н заміняються дуже тонкими, легко розтяжними нитками S невідомого хімічного складу. Комплекс, що утворюється завдяки поперечним зв'язкам між філаментами актину й міозину, називається актоміозином.
Так само, як поширюється збудження, процес скорочення підкоряється закону «все або нічого». Це означає, що при понадграничному подразненні амплітуда скорочення волокна завжди однакова, а якщо подразнення не досягає порога, вона дорівнює нулю. Однак це відноситься лише до окремого волокна, але не до всього поперечносмугастого кістякового м'яза.
Кістякова мускулатура хребетних тварин зазвичай увесь час перебуває у стані деякої напруги, що підтримується нервовими імпульсами від рухових нейронів (тонус). Гладка мускулатура може довго залишатися у скороченому стані зовсім або майже без підвищення інтенсивності свого метаболізму. У м'язових клітинах хімічна енергія перетворюється у механічну роботу. Коефіцієнт корисної дії становить 30-35%, тобто близько 2/3 енергії втрачається у вигляді тепла.
Під час скорочення м'язової фібріли актинові філаменти всуваються між міозиновими (рис. 9.5, В) і в результаті саркомери коротшають. За цей «механізм ковзання» відповідальні «голівки», наявні на нитках міозину.
9.3 Поведінка
Дослідження поведінки природно науковими методами становить завдання етології (науки про поведінку).
9.3.1 Уроджені форми поведінки
Поведінка характеризується, насамперед, рухами, що безпосередньо спостерігаються. Вона може бути уроджена (спадкоємна). Тоді вона проявляється однаковим чином у всіх представників даного виду одного віку і однієї статті, навіть якщо вони були вирощені у повній ізоляції. Такі уроджені форми поведінки називають інстинктивними рухами або спадкоємними координаціями. Сюди відносяться досить різні типи поведінки, від простих рефлекторних актів до надзвичайно складних комплексів дій (рис. 9.7). Уроджені форми поведінки можуть проявлятися відразу після народження або вилуплення (наприклад, клювання зерен, питво і розкопування землі у курчат) або пізніше (наприклад, поведінка, пов'язана з розмноженням). Вона може зникати з ходом розвитку, як, наприклад, «рефлекс чіпляння» у недоношених дитинчат (у дитинчат людиноподібних мавп він не зникає).
 
| 
|
| А | Б |
Рис. 9.7. Батьківський інстинкт (турбота про потомство).
А. Самка сумчастої квакші виношує ікру у сумці на спині. Б. Материнський інстинкт у єнота-полоскуна
Деякі вроджені форми поведінки можна викликати, пред'являючи макети природних стимулів-об'єктів. При цьому центральна нервова система тварини виділяє діючі стимули – сигнальні (ключові) подразники – з великої інформації, що пропонуєтьсяї тварині. Наприклад, самець колюшки під час охорони своєї ділянки майже не звертає уваги на точні за формою і кольором макети самців того ж виду, якщо черевце в них не пофарбовано в червоний колір. Але сильно спрощені макети з червоною нижньою стороною піддаються інтенсивним атакам (рис. 9.8).

| |
| А | Б |
Рис. 9.8. Досліди з макетами колюшки. А. Реалістичний за формою й розцвіченням макет без червоного черевця майже не викликає нападів.
Б. Спрощені макети із червоним черевцем викликають бурхливі атаки
Багато форм поведінки дуже складні, і при більш докладному аналізі з'ясовується, що це ланцюги окремих дій, причому для кожної із цих дій потрібен свій ключовий подразник.
9.3.2 Внутрішні умови та фактори
Якщо при однакових зовнішніх умовах у різний час пред'являти тій самій тварині певний стимул, то здатність його викликати реакцію часто виявляється досить різною. Для зозулі яйця або вже пташенята, що вилупилися, прийомних батьків служать ефективним стимулом тільки в перші дні життя. Вона викидає їх із гнізда. Однак на муляжі яєць, підкладених у гніздо пізніше, вона не звертає уваги.
Багато експериментів підтверджує значення гормонів як внутрішніх факторів, що визначають поведінку. Наприклад, введення пролактина будить у голубів інстинкт турботи за потомство, а у курок-несучок викликає кудкудакання. Програми поведінки можна також запускати, активуючи певні нервові центри.
Поведінкові акти звичайно не бувають ані чистими реакціями, ані чисто спонтанними діями. Як правило, вони залежать як від зовнішніх стимулів (пускових факторів), так і від внутрішніх спонукань (факторів, «що настроюють»). У разі довгої відсутності стимулюючої ситуації внутрішнє спонукання іноді може стати таким сильним, що пов'язана з ним форма поведінки може виявитися «сама по собі». Це називають «реакцією у порожнечі». Наприклад, птаха можуть почати насидження у порожнім гнізді або у собак виникає «несправжня» щенність.
Якщо понадсильне збудження не може розрядитися звичайним шляхом, можуть з'явитися «зміщені» дії (зміщена активність), що зовсім не мають відношення до даної ситуації. Так, півні можуть раптово ненадовго перервати свій бій і зайнятися клюванням чогось на землі, начебто вони голодні, шпаки в розпал бійки раптом починають чистити пір'я тощо.
9.3.3 Придбана поведінка
Уроджені інстинктивні дії можуть бути тимчасово, або назавжди змінені під впливом навчання; це будуть так називані уроджені механізми, доповнені досвідом. При цьому може змінитися характер реакції або ж механізм її запуску. Різні форми поведінки у тієї самої тварини можуть в різному ступені піддаватися зміні у результаті навчання.
Навчання шляхом наслідування може приводити до утворення місцевих традицій. Відомо, наприклад, що у пацюків уникнення отруєної принади, з'явившись у декількох особин, може поширюватися на всю зграю й зберігатися у ряді поколінь, якщо приманка розкладується регулярно.
Ворони вчаться розрізняти придатний і менш придатний для будівлі гнізда матеріал методом проб і помилок, причому спочатку вони тягнуть до гнізда все, що можна, не перебираючи. Ссавці в такий же спосіб навчаються вибирати більше гострі краї замість рівних поверхонь, коли хочуть почухатися об щось.
Найвища ступінь придбаної поведінки – це поведінка, яка заснована на розумінні (інсайті); вона зустрічається тільки в самих високорозвинених представників хребетних.
Інсайт стає особливо помітним під час планомірнихдійта використанніпредметів. Щоб дотягтися до високо підвішеного банана, шимпанзе можуть скріплювати палки різної товщини або нагромаджувати один на одного ящики. Але для цього потрібно, щоб тварини раніше могли придбати деякий досвід використання даних предметів. Виготовлення знарядь відомо і у шимпанзе, що живуть на волі.
9.3.4 Орієнтація у просторі
У сидячих тварин (коралові поліпи та ін.), а також у рослин відомі тропізми, які обумовлені та направлені ростовими реакціями. У рослин фототропізм і геотропізм – реакція на земне тяжіння (силу ваги); негативна у стебла та позитивна у кореня. Наприклад, надламане стебло трав'янистої рослини, викривляючись, продовжує рости вертикально; при вирощуванні проростка в трубці, його стебло й корінь ростуть горизонтально, але як тільки вони вийдуть із кінця трубки, що обмежує ріст, стебло згинається до верху, а корінь – униз (рис. 9.9).
Настії – рух рослин, що викликаються подразненням, але вони не мають відношення до орієнтованого руху, тому що їхня спрямованість визначається структурою реагуючого органа. Приклади: сейсмонастія; відкривання й закривання багатьох квіток під час зміни світла й темряви (фотонастія, наприклад, у латаття) або зміні температури (термонастія, наприклад, у тюльпана).
Рис. 9.9. Геотропізм рослин
У організмів, що вільно пересуваються (бактерій, джгутиконосних водоростей, тварин) і репродуктивних клітин (спір та гамет, що мають джгутики) розрізняють кінези й таксиси. Під час кінезів рух буває ненаправленим, однак при наявності градієнта подразника організми, зрештою, збираються в найбільш підходящій для них зоні.
Таксиси – це спрямовані переміщення до джерела стимуляції (позитивні таксиси) або від нього (негативні таксиси). При клинотаксисі курс визначається шляхом «обмацування» середовища, наприклад маятникоподібним рухом переднього кінця тіла або обертанням усього тіла. Телотаксис заснований на прямої фіксації мети.
Під менотаксисом розуміють вибір твариною курсу під якимсь кутом від 00 до 1800 до напрямку на джерело подразника. Наприклад, багато членистоногих та хребетних орієнтуються за світлом, але рухаються не прямо до джерела світла або від нього, а підтримують під час свого руху певний кут до напрямку сонячних променів (орієнтація за сонячним компасом).). Багато тварин, що орієнтуються шляхом фотоменотаксису, дивно точно враховують добовий рух Сонця; нерідко вони здатні зберігати менотаксичний курс навіть тоді, коли можуть бачити не саме Сонце, а лише ділянка блакитного піднебесся: вони можуть визначати положення Сонця за зміною поляризації розсіяного світла піднебесся.
При орієнтації в індивідуальному життєвому просторі (територіальній орієнтації) часто велику роль грає запам'ятовування примітних орієнтирів (у ос, бджіл, мурах, птахів та ін.).
В експериментах з перевезенням тварин у ряді видів була виявлена надзвичайно розвинена здатність до знаходження будинку. Шпаки поверталися до своєї гніздової ділянки з відстані 465 км, сільські ластівки – з відстані 1800 км, а буревісники – навіть до 5000 км. Щоб визначати своє місцезнаходження птах повинен мати справжні навігаційні здібності. Механізми, що лежать в їхній основі все ще не цілком зрозумілі. У разі вибору напрямку поштові голуби використовують «магнітний компас», а також згаданий вище «сонячний компас». Такі методи орієнтації, безсумнівно, використовуються і під час сезонних міграцій. У багатьох птахів вірогідно продемонстрована також орієнтація за зірковим компасом.
9.3.5 Біокомунікація
Форми обміну інформацією (комунікації) між тваринами різноманітні. Передані сигнали можуть мати хімічну, оптичну, електричну або механічну природу.
Хімічна сигналізація – найпоширеніший і, можливо, самий древній спосіб. Речовини, що впливають на рецептори та служать для обміну інформацією між особинами одного виду, називають феромонами. Сюди відносяться статеві атрактанти (наприклад, у нічних метеликів), речовини для того, щоб позначати територію або для прокладання пахучих слідів (наприклад, «мурашиних доріг»), а також феромони тривоги, що викликають реакції страху та втечі (у багатьох прісноводних рослинноїдних риб) або підвищену агресивність (у мурах і бджіл) в особин того ж виду.
При оптичній сигналізації можуть використовуватися кольори і форми у вигляді постійних або короткочасних сигналів. Постійні сигнали (кольори або форми) служать для повідомлення про видову належність, стать, а часто і індивідуальні особливості; форми або кольори, що демонструються короткочасно, повідомляють про певні стани, наприклад про стан статевої активності (шлюбне вбрання у риб і птахів), про загальне збудження або готовність до ворожих дій. Збільшення силуету тіла, його випрямлення (у кішки – згорблення), здиблення шорстки, куйовдження пір'я, розпрямлення в різні сторони кінцівок або інших придатків тіла – типово загрозливіжести. Часто ці жести супроводжуються звуковими сигналами (фиркання, ричання тощо) і характерними рухами. Жести підпорядкування (пози покірності), навпроти зазвичай пов'язані зі зменшенням силуету тіла (скорчена поза).
Механічна сигналізація може виконуватись шляхом тактильних вібраційних або звукових стимулів, при цьому останні – філогенетично наймолодіші. Передача повідомлень за допомогою звуків зустрічається у комах та хребетних. Співочі птахи, леви та морські леви акустично "позначають" свої ділянки. У самців комах (коників, цвіркунів, цикад тощо), а з хребетних – насамперед у птахів, відомі закличні шлюбні сигнали. У птахів та ссавців звукова сигналізація, крім того, відіграє велику роль під час вирощування потомства. У звукових сигналів є та перевага, що вони ефективні і у разі відсутності зорового контакту або у темряві; з іншого боку, їх можуть вловлювати і ті тварини, якім вони не адресовані, так що ті тварини, що сигналізують можуть бути знайдені ворогами. Ще одна перевага акустичних сигналів – вони можуть бути у більшій ступені структуровані у часі і таким чином дозволяють передавати більше інформації.
Навіть вищі форми голосової комунікації у тварин принципово відрізняються від мови людини, яка засвоюється шляхом навчання. Повідомлення тварин завжди стосуються обставин, важливих на цей момент. Тварини не спроможні повідомляти про події, що вже закінчились або тільки очікуються у майбутньому. Тільки людини спроможна за допомогою своєї мови «абстрактно мислити словами, створювати вищі відвернені поняття, виражати причинні та логічні відносини і, насамперед, створювати традиції».
Запитання для самоперевірки
1. Основні рецептори та будова рецепторних клітин.
2. Потенціал спокою та його хімічні характеристики
3. Потенціал дії та його хімічні характеристики.
4. Проведення збудження міелінізованими та неміелінізованими нервовими волокнами.
5. Синаптична передача збудження.
6. Рефлекторна дуга та рефлекс.
7. Типи процесів навчання.
8. Форми пам’яті, що виділяють різні автори.
9. Рух та його типи.
10. Особливості м’язового руху.
11. Уроджені форми поведінки.
12. Придбана поведінка.
13. Тропізми, настії, кінези та таксиси.
14. ”Сонячний, магнітний та зірковий компаси”.
15. Хімічна, оптична та механічна сигналізації у тварин.
Лекція 15
ГЛАВА 10
|
|
|
|
|
Дата добавления: 2015-05-24; Просмотров: 1786; Нарушение авторских прав?; Мы поможем в написании вашей работы!