КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Белоктарды топтастыру. 1 страница
|
|
|
|
Белок молекуласы пішіні жағынан екі топқа бөлінеді: глобулярлы (шар немесе элипс тәрізді); фибрилярлы (талшық тәрізді) белоктар. Глобула яғни шар терізді белоктарда бір немесе бірнеше полипептидтік тізбектер клетка ішіндегі кеңістікте тығыз құңдақталып, жиырылып шар тәрізді құрылым береді. Олар суда ериді. Бұларға көптеген белоктар, солардың ішінде ферменттер жатады. Талшық тәрізді белоктар суда ерімейді молекуласы ұзын жіп тәрізді, полипептидтік тізбек бір осьтің бойында созылып орналасқан. Көпшілік талшық тәрізді белоктар кұрылымдық және қорғаныштық қызмет атқарады. Олардың нағыз өкілдері шаштағы жүндегі α-кератин, жібектегі фибрион және сіңірдегі коллаген. Осыңдай белоктар қатарына бұлшық еттің жиырылып созылу жүйесін қамтамасыз ететін актин, миозин жатады.
Осы екі топқа жататын белоктардың бір-бірінен айырмашылығы, олардың құрамындағы аминқышқылдарына байланысты. Талшықты белоктардың құрамында гидрофобты аминқышқылдары басым (фенилаланин, изолейцин, валин, метионин, тирозин, триптофан) және олардың радикалдары бір-бірімен оралып жатқан тығыз тізбектін, сырт жағынан орналасқан, олардың суда ерімейтіні негізінен осыған байланысты. Глобулярлы, шар тәрізді белоктар да радикалы гидрофобты аминқышқылдары көп болуы мүмкін, бірақ олардың полипептидті тізбегі қомақталып жинақталған кезде олар глобуланың ішкі жағында орналасады да, сумен жақындаспайды.
Химиялық табиғатына қарай, жоғарыда айтылғандай, белоктар жәй қарапайым-протеиндер, және күрделі - протеидтер болып екіге бөлінеді.
Қарапайым белоктар тек амин қышқылы қалдықтарынан тұрады, ал күрделі белоктардың құрамында белоктың бөлігінен басқа жанама табиғаты белок емес бөлігі болады.
Осы екі топ өзара топшаларға яғни шағын топтарға бөлінеді. Протеиндерді әртүрлі ертінділерде ерігіштігіне байланысты топқа бөледі.
1. Альбуминдер – бейтарап тұздармен, мысалы, аммоний сульфатымен немесе натрий сульфатымен толық қанықтырған кезде тұнбаға түседі. Протеиндердің өкілдері суда еритін альбуминдер, табиғатта кеңінен тараған белок. Әсіресе адамның қан плазмасының құрамында көп, оның үлесіне плазмадағы жалпы белок мөлшеріне шаққанда 50% тиеді. Альбумин молекуласы теріс зарядталған. Құрамында глутамин қышқылы болғандықтан, оның қышқылдық қасиеті бар. Екінші бір кең тараған өкілі глобулин. Глобулиндер көптеген өсімдіктердің дәніндегі белоктың негізін құрайды, әсіресе бұршақ тұқымдас, майлы өсімдіктердің дәнінде көп болады. Глобулин сәл қышқылдық қасиеті бар немесе бейтарап белоктарға жатады. рН = 6 – 7,3. глобулин қанда көп кездеседі.
60-80%-ті этил спиртінің ерітіндісінде еритін қарапайым белок- проламиндер.
Бұлар тек қана астық тұқымдас өсімдіктердің дәнінде кездеседі, онда олар қор ретінде жиналады. Бидай мен қара бидай дәнінде –глиадин деген белок, ал арпа дәнінде – гордеин, жүгеріде – зеин деген спиртте еритін белоктар мөлшері өте көп болады. Бұл белоктар тобында ауыстырылмайтын амин қышқылдары тапшы болады.
Сілті ерітіндісінде (0,2-2%NaOH) еритін белоктар – глютелиндер. Бұларда өсімдіктерде тараған, олардың дәнінде біраз деңгейде болады. Бидай дәнінде болатын осы топшаға жататын глутелин деген, ал жүгеріде – оризенин деген белоктар бар.
Протаминдер – ең шағын молекулалы белоктар, М.М. – 12000шамасындай, күшті негіздік қасиеті бар, құрамында аргинин қалдығы көп, гистондар сияқты ДНҚ – мен байланысты. Ол спермада (еркектің ұрығында) болады және реттеушілік емес, тек құрылымдық қызмет атқарады.
Сілтілік қасиет көрсететін белоктар - гистондар. Олар әлсіз қышқылдарда (0,2 немесе HC1) ериді, аммиак пен спиртпен тұнбаға түседі.
Негізінен бұл белоктар жануарлар мен өсімдіктер жасушаларының ядроларында кездеседі және хроматин құрылымында маңызды рөл атқарады. ДНҚ – ның кеңістіктік құрылымынтұрақтандыруға қатысады және генетикалық мәліметтің ДНҚ – дан РНҚ – ға берілуіне жан-жақты тосқауыл қояды.
Күрделі белоктар құрамындағы жанама тобының табиғатына қарай бірнеше топшаларға бөледі. Олар: липопротеидтер, фосфопротеидтер, гликопротеидтер, хромопротеидтер, нуклеопротеидтер, металлопротеидтер.
Литопротеидтер табиғатта көп тараған, барлық тірі ағзаларда кездеседі. Олар барлық жасуша мембраналарының ең бір басты құрамы болып есептелінеді. Липопротеидтер бос күйінде қан плазмасында және лимфада кездеседі және ол липидтерді, кейбір гормондарды бір орнынан екінші орынға ауыстырып апара алады. Электрофорездік қозғалғыштығы бойынша α – липопротеидтерге және β – липопротеидтерге бөлінеді. Фосфопротеидтер құрамында ортофосфор қышқылы бар, олардың ағзаның қоректенуінде маңызы зор. Сүт құрамындағы -казеин, жұмыртқа сарысындағы – виттелин, балық уылдырығындағы – ихтулин осы фосфопротеидтерге жатады. Гликопротеидтер құрамында белок емес бөлігі – көмірсулар. Көмірсулардан олардың құрамына кіретіндер нешетүрлі моносахаридтер, дисахаридтер, тіптен полисахаридтер де бола алады.
Гликопротеидтер жануарларда, өсімдіктерде, микроорганизмдерде кездеседі және олар әртүрлі қызметтер каталитикалық, тасымалдаушы т.б.атқарады.
Хромопротеидтер – боялған күрделі белоктар, олардың құрамында бояулы заттар болады. Олардың нағыз өкілдері родопсин, гемоглобин, миоглобин, пероксидаза, каталаза, флавиндік ферменттер және цитохром жүйесіне жататын ферменттер.
Нуклепротеидтердегі жанама топ нуклеин қышқылдары. Нуклеопротеидтердің молекулалық массасы үлкен болып келеді. Дезоксинуклеопротеидтер жасушаның ядросында, митохондрияларында кездеседі, ал рибонуклеопротеидтерге – рибосомалар мен информосомаларды жатқызуға болады. Вирустарда да өзінің химиялық табиғаты жағынан – нуклеопротеидтер. Нуклеопротеидтердің белок биосинтезінде және тұқым қуалаушы информацияны сақтауда маңызы зор.
№5 дәріс
Тақырыбы: Матрицалық (қалыптық) биосинтез
Жоспар: 1.ДНҚ репликациясы
2. ДНҚ молекуласының негізгі бөлімінің репликациялануы
3. Репликация тетіктері
ДНҚ молекуласының ең маңызды қасиеттерінің бірі — оның өздігінен екі еселенуі (репликациялануы) болып саналады. ДНҚ репликациялануы салдарынан тұқым қуалаушылық ақпарат ұрпақтан - ұрпаққа өзгеріссіз, тепе-тең мөлшерде беріліп, ұрпақтардың жалғасуы қамтамасыз етіледі. ДНҚ репликациясы жасуша циклының S -синтетикалық кезеңінде жүзеге асады.
ДНҚ молекуласының репликациялану қасиеті 1953ж. Дж. Уотсон және Ф.Криктің ДНҚ молекуласының құрылысының қос ширатпалы болатындығын анықтағаннан кейін белгілі болды.
Теория күйінде ДНҚ репликациясының 3 түрлі әдісі болжамдалған: 1) консервативті (тұрақты); 2) жартылай консервативті; 3) дисперсті.
Көптеген тәжірибелер нәтижесінде ДНҚ молекуласының репликациялануы жартылай консервативті жолмен жүретіндігі дәлелденді. Оны алғашқылардың бірі болып 1958ж. М.Мезельсон және Ф.Сталь Е.соІі жасушасында байқаған.
Қазіргі таңда ДНҚ молекуласының сырт пішінінің 3 түрі белгілі: тұрақты сақиналы (бактериофакторда); құбылмалы сақиналы (бактериофактарда); сызықты (прокариоттар және эукариоттарда). Осыған сәйкес ДНҚ молекуласының жартылай консервативті репликациялануының 3 түрі белгілі: 1) тета репликация; 2) сигма репликация; 3) У-тәрізді репликация.
Кейбір прокариоттардың және барлық эукариоттардың ДНҚ молекуласы сызықша тәрізді болып келеді және олардың репликациялануы белгілі бір нүктеден, репликативтік ісінудің пайда болуынан басталып, хромосоманың қарама-қарсы жағына қарай бағытталады. Эукариоттардың ірі хромосомаларында бір мезгілде жүздеген репликациялық ісінулер пайда болады және олар бір — бірімен қосылып У-тәрізді аралық құрылым пайда етеді. Мұны У-тәрізді жартылай консервативті репликациялану деп атайды.
ДНҚ репликациясының бірнеше ерекшеліктері белгілі:
а) ДНҚ молекуласының жаңа тізбегінің синтезделуіне қажет заттар — дезоксинуклеозидтрифосфаттар (дНТФ) болып табылады, ал ДНҚ құрамында дезоксинуклеозидмонофосфаттар (дНМФ) кездеседі. Сондықтанда ДНҚ тізбегіне жалғану алдында әрбір нуклеотидтен 2 фосфат қалдығы пирофосфат күйінде бөлініп шығады да тез арада фосфаттарға дейін гидролизденеді.
Еркін дНТФ —> дНМФ қалдығы + пирофосфат
дНТФ-ды құрылыс материалдары ретінде пайдаланудың энергетикалық себептері де бар. Нуклеотидтер арасындағы байланыстардың (фосфодиэфирлік) түзілуі үшін энергия қажет, ал энергия фосфаттараралық байланыстардың үзілуі нәтижееінде бөлінеді.
б) ДНҚ репликациясы матрицалық (қалыптық) үдеріс яғни ДНҚ- ның жаңа тізбегі аналық ДНҚ молекуласының бір жіпшесі негізінде
(матрица) комплиментарлық ұстаныммен (принциппен) синтезделінеді,
яғни 4 нуклеотидтен (дАТФ, дГТФ, дЦТФ, дТТФ) жаңа тізбекке
тек аналық жіпшедегі нуклеотидке комплиментарлы (А<->Т; Г<-Ц)
нуклеотид қана қосылады.
в) ДНҚ синтезі (репликациясы) симметриялы болады, яғни матрица
қызметін аналық ДНҚ молекуласының екі тізбегі де атқара береді.
Сондықтан оны жартылай консервативті деп те атайды. Себебі,
жаңадан синтезделген ДНҚ молекуласы жартылай жаңарған болады,
яғни оның бір тізбегі ескі -аналық молекуладан алынған болса
(матрица), екіншісі жаңадан синтезделген болады.
г) ДНҚ синтезі (жаңа тізбектің не оның бір бөлімінің синтезделуі)
белгілі бір бағытта жүреді, яғни 5' ұшынан 31 ұшына карай жүреді.
д) ДНҚ репликациясы басталу, жүруі үшін, міндетті түрде
аналық ДНҚ молекуласының қос ширатпасы бір бірінен ажырасуы
қажет, тек осы жағдайда, яғни бір бірінен ажырасқан аналық
молекуланың жіпшелері матрица (қалып) қызметін атқара алады
Репликация тетіктері
а) Репликация үдерісі 15-20 ақуыздардан тұратын күрделі ферменттік
жүйенің қатынасуымен жүзеге асады.
Эукариоттар хромосомаларында жоғарыда айтқанымыздай, бір мезгілде көптеген ферменттік кешендер қызмет етеді, яғни хромосомада ДНҚ репликациясының көптеген басталу (инициация) нүктелері болады және ДНҚ синтезі хромосоманың бас жағынан ұшына қарай баяу жүрмей, көптеген жерлерінде бір мезгілде жүзеге асады. Бұл репликация ұзақтығын едәуір қысқартады.
б) репликацияның әр бір нүктесінде 2 ферменттік кешен жұмыс
істейді: олар ДНҚ-ның инициация нүктесінен қарама-қарсы бағыттарға
қарай жүреді.
в) ДНҚ молекуласының тізбектері бір-біріне антипаралель
болғандықтан және ДНҚ синтезі тек 51—>-3' бағытында жүретіндіктен,
репликативтік ашада аналық ДНҚ-ның бір тізбегі негізінде жаңа
ДНҚ тізбегі үзіліссіз синтезделсе, екіншісі негізінде үзіліп-үзіліп
синтезделінеді. Біріншісін лидерлік тізбек, ал екіншісін артта қалушы
(кешігуші) жіпше деп атайды.
Лидерлік тізбек негізінде синтезделген өте ұзын, ұзындығы көршілес екі инициация нүктелерінің ұзындығының жартысына, яғни 1.600.000 нуклеотидке тең, тізбек синтезделсе, артта қалушы (кешігуші) тізбек негізінде қысқа 1500 нуклеотидтерден тұратын ДНҚ фрагменттері синтезделінеді. Оларды Оказаки фрагменттері деп атайды.
г) ДНҚ синтезі басталуы үшін міндетті түрде 10-15 нуклеотидтерден
тұратын «РНҚ-ұйытқы-праймер қажет, себебі ДНҚ синтезін жүргізетін
фермент ДНҚ - полимераза өз бетінше ДНҚ синтезін бастай
алмайды.
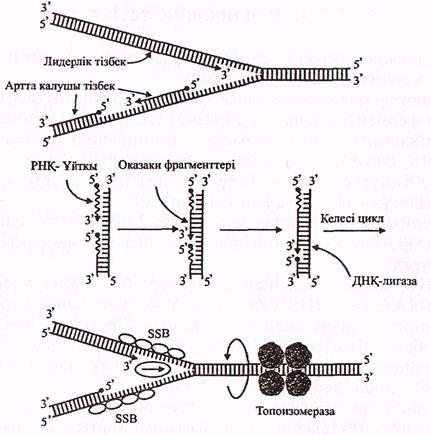
1-сурет. ДНҚ репликациясының жобасы (Айала, Кайгерден, 1987)
1. Аналық ДНҚ молекуласын репликациялауға дайындайтын ақуыздар.
а) ДНҚ молекуласындағы репликацияның басталу (инициация) нүктелері А-Т нуклеотидтер жұптарына бай бірізділіктерге ие. Ерекше танып білуші ақуыз (А) әрбір осындай нуклеотидтер бірізділігіне (А-Т бай) ДНҚ-репликациялаушы кешенді байланыстырады да өзі әрі қарай, кешенмен бірге жылжымайды (2-сурет).

2-сурет. ДНҚ-репликациялаушы кешен (Мушкамбаров, Кузнецовтан, 2003)
А-танып білуші ақуыз; Г-геликаза; И-топоизомераза; S-SSВ-ақуыз; П-праймаза; АП-праймаза активаторы; α,β,γ-ДНҚ полимеразалар; Р-РСNА-ақуыз; Н-нуклеаза; ДНҚ лигаза (Мушкамбаров, Кузнецовтан, 2003).
б) Ферменттік кешеннің ең алғашқы іске кірісетін ферменті-
геликаза (Г). Ол аналық ДНҚ молекуласының қос ширатпасының
ашылып, жіпшелердің бір-бірінен ажырасуын қамтамасыз етеді.
в) Ширатпаның ашылып жазылуы әрі қарай үлкенді-кішілі
түйіндердің (суперспирализация) пайда болуына алып келеді. Мұның
себебі әрбір ДНҚ молекуласы өздерінің бірнеше учаскелерімен ядро
матриксіне бекінген, ал бұл ширатпалары ашылған ДНҚ
молекуласының еркін айналуына мүмкіндік бермейді, сондықтан да
түйіндер пайда болады.
Бұл мәесле топоизомераза (И) ферментінің қатынасуы арқылы шешіледі (3-сурет).
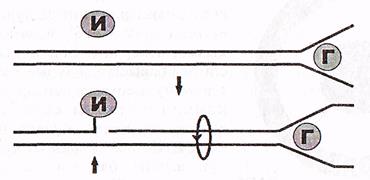
3-сурет. Топоизомераза І-әрекеттері (Мушкамбаров, Кузнецовтан, 2003)
г) Сонымен, геликаза, топоизомераза ферменттері аналық ДНҚ молекуласының қос ширатпасын жеке-жеке екі тізбекке ажыратады. Ажырасқан әрбір тізбекпен ерекше SSВ-ақуыздар байланысады, олардың қызметі бір-бірінен ажырасқан жіпшелерді керіп, күні бұрын жанасып, қос ширатпаның түзілуін болдырмау болып табылады.
№6 дәріс
Тақырыбы: Полимеризация ферменттері
Жоспар: 1.ДНҚ репликациясын аяқтаушы ферменттер
2. ДНҚ-ның теломерлік бөлімдерінің репликациялануы
3. Теломералар қызметтері
а) ерекше ақуыз праймаза активаторы (АП) қызметін атқарады.
Осыдан кейін праймаза (П), бір жіпшелі ДНҚ-ның тиесілі учаскесін
матрица (қалып) ретінде пайдаланып қысқа «РНҚ-ұйытқыны» (праймер)
синтездейді.
б) Әрі қарай ДНҚ синтезін ДНҚ, полимераза жүргізеді.
Эукариоттарда 5 түрлі ДНҚ полимеразалар белгілі: β және ε
полимеразалар ДНҚ репарациясына қатынасады; γ-полимераза — митохондрия ДНҚ-сының репликаңиясын жүзеге асырады; ал α және σ полимеразалар — ядролық ДНҚ репликациясына қатынасады.
α ДНҚ полимераза праймазамен де, σ —полимеразамен де байланысады, ал соңғысы РСNА (Р) деп аталатын ақуызбен байланысқан.
РСNА (Р) ақуыз-полимераза кешенін ДНҚ-ның репликацияланушы тізбегіне бекіндіріп «қыстырғыш» рөлін атқарады. РСNА ақуыз «қыстырылған» күйінде сақина сияқты ДНҚ тізбегін қоршап тұрады және полимеразалардың ДНҚ тізбегінен күні бұрын диссоциациялануын (ажырауын) болдырмайды, яғни ДНҚ синтезінің жүруіне, жалғасуына мүмкіндік жасайды (4-сурет).

РСNА ақуызымен ДНҚ тізбегінің әрекеттесуі
ДНҚ—полимеразалар (әсіресе бактериялардың ДНҚ-полимераза ІІІ-) нуклеотидтердің аналық тізбекке комплиментарлы синтезделуін қамтамасыз етуімен бірге 31—>5'-экзонуклеазалық-та қызмет атқарады. Соңғы қызметі ДНҚ синтезі барысында дұрыс —комплиментарлы нуклеотидтердің орнына, бұрыс, комплиментарлы емес нуклеотид, жалғанған кезде жүзеге асады. Осы кезде ДНҚ-полимераза бұрыс жұптасқан нуклеотидті «байқап» қалып, оны өсіп келе жатқан 31 ұшынан шығарып (үзіп)
алып тастайды. Осылайша полимеразалар өз жүмыстарын үнемі бақылап отырады
в) Кез келген жаңадан еинтезделген ДНҚ фрагменттері-(ұзын лидерлік не
Оказаки фрагменттері) праймерлерден («РНҚ-ұйытқыдан) басталады. Аналық (матрицалық) тізбек бойымен жылжып отыратын ферменттік кешен келесі ДНҚ фрагментіне жанасқаннан кейін, ДНҚ-полимераза ІІІ ферменті «қыстырушы» РСNА ақуызын ашып, кешенді матрицадан ажыратады және ДНҚ синтезін тоқтатады.
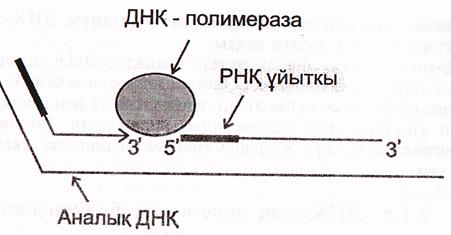
5-сурет синтезделген ДНҚ фрагменттерінің түйісуі (Мушкамбаров, Кузнецовтан, 2003)
Осыдан кейін ДНҚ полимераза І-іске кіріседі. Ол өсіп келе жатқан ДНҚ фрагментінің 3' ұшына жалғанады және 3 белсенділікке ие болады.
Біріншіден ол «алдыңғы» немесе 51—>3'-экзонуклеазалық белсенділікке ие болады, яғни ол бұрынғы ДНҚ тізбегінің «РНҚ-ұйытқысының» (праймер) 51 ұшынан бір-бірлеп нуклеотидтерді алып тастап отырады, ал босаған жерге өз фрагментінің 31 ұшына дезоксинуклеотидтерді жалғайды (ДНҚ-полимеразалық белсенділік).
Сонымен қатар, ДНҚ-полимераза ІІІ-сияқты «артқы» 31—>5' экзонуклеазалық белсенділік арқылы өз жұмысын қадағалауды да «ұмытпайды».
ДНҚ-полимераза-І-қызметі өсіп келе жатқан ДНҚ фрагментінің бұрынғы ДНҚ фрагменттерінің дезоксинуклеотидтерімен түйіскеннен кейін аяқталады.
Эукариоттарда ДНҚ-полимераза ІІІ-қызметін α және σ—ДНҚ полимераза кешені атқарады; бұл жерде 31—5' экзонуклеазалық белсенділік σ -ДНҚ —полимеразаға тиесілі болса, ДНҚ-полимераза І-қызметін, 51—>3' —экзонуклеазалық қызметті ерекше фермент нуклеаза (Н), ДНҚ-полимеразалық белсенділікті (босаған жерді толтыру) β -ДНҚ - полимераза атқарады.
Жоғарыда аталған ферменттер кешені қызметтері нәтижееінде жаңадан синтезделінген әрбір тізбектер бір-бірімен тығыз орналасқан көптеген фрагменттерден тұрады.
Көршілес фрагменттердің бір-біріне жалғануы ДНҚ-лигаза (Л) ферменттері арқылы жүзеге асады.
ДНҚ-полимераза ферменттері сияқты ДНҚ-лигазалар-да нуклеотидтерді фосфодиэфирлік байланыс арқылы байланыстырады. Осылайша ДНҚ молекуласының негізгі бөлігі репликацияланады, ал оның ұштары, яғни теломералық учаскелері, ерекше жолмен репликацияланады. Бұл үдеріске ерекше ферменттер теломеразалар қатынасады.
ДНҚ молекуласының толық репликацияланбайтындығын, яғни теломерлік бөлімдерінің репликацияланбайтындығын, алғаш рет 1971ж. А.М. Оловников айтқан болатын.
Мұның мәні мынада: жоғарыда сипатталған ДНҚ полимеразалық жүйе аналық ДНҚ молекуласының жіпшелерінің 31 ұшын толық репликацияламайды, яғни жаңадан синтезделген ДНҚ тізбектері 51 ұшы жағынан қысқа болады. Себебі әрбір жаңа ДНҚ тізбегі қысқа «РНҚ – ұйытқыдан» (праймер) басталады. Кейін ол ерекше нуклеазалар арқылы алынып тасталады, бірақ босаған учаске дезоксинуклеотидтермен толтырыла алмайды, себебі ДНҚ полимеразалар өз бетінше («РНҚ-үйытқысыз») ДНҚ синтезін бастай алмайды, ол тек полинуклеотидті З1 ұшынан ұзартады. Бұл жерде ондай учаске жоқ, сондықтан жаңа тізбек матрицадан қысқа болады.
ДНҚ молекуласының мұндай ұшын (бір тізбегі ұзын, екіншісі қысқа) үшкір ұшы немесе оверхенга деп аталады.
ДНҚ-ның үшкір үшы тұрақсыз болады, себебі экзонуклеазалар ұзын ұшындағы артық нуклеотидтерді бір-бірлеп алып тастап, ДНҚ ұшын тұйықтайды.
Қалай болғанда да, егер жасушада теломераза болмаса, оның әрбір бөлінуінен кейін хромосома (ДНҚ) қысқарып отырады.
Әрбір репликацияда ДНҚ молекуласы «РНҚ – ұйытқы» ұзындығына сәйкес 10-15 нуклеотидке қысқаруы тиіс болғанымен, шындығында 50-65 нуклеотид жұбына қысқарады. Бұл ДНҚ-полимеразалық кешеннің қасиетіне байланысты болады.
Адамның ядролық ДНҚ-ның 1 молекуласының орташа ұзындығы 120 миллион нуклеотид жұптарына тең десек, жасушаның әрбір бөлінуінде теломераза белсенділігінсіз ДНҚ молекуласы 0,00005%-ға қысқарады екен. Бұл әрине өте аз. Бірақ, табиғатта теломера
ұзындығын қалпына келтіріп отыратын тетіктер болмаса түбінде хромосомалар жойылып кеткен болар еді. Тек сондықтан ғана хромосомалар теломерлерінің толық репликацияланбау проблемасының биологиялық маңызы орасан зор. Сонымен қатар, бұл құбылыс ағзалардың қартаю, канцерогенез проблемаларымен де тығыз байланысты.
Толық репликацияланбау проблемалары жасушада қалай шешіледі?
Ғылыми деректер бойынша хромосома ұштарында генетикалық ақпарат болмайтын көптеген арнайы гексонуклеотид (6 нуклеотидтен тұратын) бірізділіктер қайталанып орналасқан.
(51) ЦТААЦЦ... ЦТААЦЦ.... ГГТТАГ... ГГТТАГ... (31)
(31) ГАТТГГ... ГАТТГГ... ЦЦААТЦ... ЦЦААТЦ... (51)
ДНҚ-ның теломерлік бөлімдерінде мыңдаған осындай гексонуклетидтер қайталанады. Олардың жалпы ұзындығы адам эмбрионы жасушаларында 10-15 мың нуклеотид жұптарына тең. Сонымен, хромосоманың екі теломерлік ұшы, адамның ядролық ДНҚ молекуласының ұзындығының 0,02 құрайды.
Теломерлік қайталануларда ешқандай генетикалық ақпарат болмайды, сондықтан да теломерасыз олардың біршама бөлігі түсіп қалған күннің өзінде де геном бірқалыпты қызмет ете береді. Теломерлердің негізгі қызметінің өзі де осы болса керек, яғни олар геномның маңызды (ақпаратты) бөлімін толық репликацияланбаудан қорғап, буферлік қызмет атқарады.
Әйтсе де, теломеразалардан бір жолата бас тартуға да болмайды, себебі жасушаның бөліну үдерісінде күндердің күнінде ДНҚ-ның теломерлік учаскелері қысқарып-қысқарып жойылуы мүмкін. Сонымен қатар, теломерлік учаскелер ерекше, арнайы қызметтер де атқарады, сондықтан ол белгілі бір шекке дейін ғана қысқарады.
Теломералар қызметтері
Теломералар төмендегідей маңызды қызметтер атқарады:
1) механикалық қызметі:
а) Теломералар хромосомалардың ядро матриксіне бекінуіне қатынасады;
б) Теломералар хромосома хроматидаларының ұштарын бір-бірімен
тіркестіреді;
2) Тұрақтандырушы қызметі:
а) Жасушада теломераза болмаған жағдайларда ДНҚ-ның кодтаушы бөлімін толық репликацияланбаудан сақтайды; Егер жасушада теломераза болса, үзілген хромосома ұштарын тұрақтандырады;
3) Гендердің экспрессиялануына әсер етуі.
Теломераға жақын орналасқан гендер экспрессиясы төмен болады (репрессияланған), мұны транскрипциялық үнсіздік немесе сайлинсинг деп атайды.
Теломерлердің айтарлықтай қысқаруы оларға жақын орналасқан гендерді активтендіреді, мысалы Rар 1 не ТҒR 1 гендерінің активтенуі.
4) «Есептеу» қызметі.
ДНҚ-ның теломерлік бөлімдері теломеразасыз жасушаның бөлінуін есептеп отыратын репликометр болып табылады. Жасуша үшін оның қанша рет бөлінгеніне қарағанда, теломера ұзындығының сындарлы деңгейіне дейін қанша рет бөлініп қалғаны маңыздырақ. Сондықтан да теломера- жасушаның теломеразасыз қанша рет бөліне алатынын есептейтін құрылым болып табылады.
Теломера ұзындығы сындарлы деңгейге жеткенде ол өзінің жоғарыда аталған қызметтерін атқара алмайды, сондықтан да жасуша циклы бұзылып өледі.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 2476; Нарушение авторских прав?; Мы поможем в написании вашей работы!