КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Белоктарды топтастыру. 4 страница
|
|
|
|
Гендердің экспрессиялануының реттелу механизмдерін зерттеу үшін прокариоттар өте қолайлы объект болып саналады, себебі олардың геномдары небәрі бірнеше гендерден құралған және олар өте қарқынды көбейеді. Сонымен қатар, гендердің экспрессиялануының реттелу механизмдері прокариоттарда және эукариоттарда да ұқсас жоба күйінде жүретіндігі белгілі болды.
Бактериялардың бірнеше алмасу реакңияларын катализдейтін ферменттердің гендері оперон деп аталатын құрылымдық- қызметтік бірлікке біріктірілген.
Оперонның құрамына аталған гендермен қатар промотор және оператор да кіреді. Промотор-РНҚ-полимеразамен байланысып, ген ақпаратының көшіріліп жазылуының (транскрипциясының) басталатын нүктесі болса, оператор-ақуыз репрессормен әрекеттесетін орын. Ақуыз репрессорды-репрессор-гені кодтайды және ол оперон құрамына кірмейді
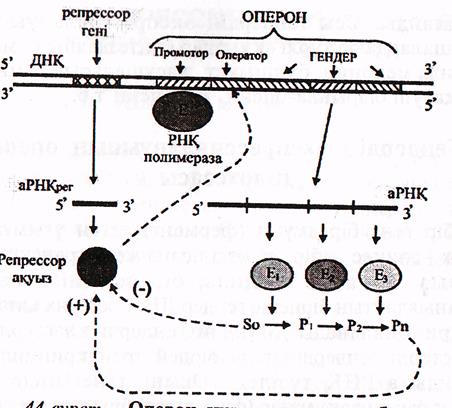
(24-сурет).. Оперон құрылысының жобасы (Мушкамбаров, Кузнецовтан, 2003)
Оперонның екі түрі белгілі.
а) Индукцияланатын оперондар:
-реттеуші болып бақылау реакциялар тізбегінің бастапқы өнімі (S о)-(лактоза, не аллалактоза) саналады-лактоза опероны; -егер ортада бұл субстрат (өнім) болмаса ақуыз-репрессор оператормен байланысып, РНҚ полимеразаның оперон гендерін транскрипциялау қызметін бастырмалайды (оперон «өшірілген»);
-егер ортада алғашқы өнім (субстрат) (S 0) болса не жинақтала бастаса, оның кейбір бөлігі ақуыз-репрессормен байланысып, оның оператормен қосылуын болдырмайды; оперон «іске қосылады» және алғашқы өнімді
(S 0) ыдыратушы ферменттер синтезделінеді.
б) Репрессияланатын оперон:
-реттеуші болып бақылау реакциялар тізбегінің акырғы өнімі (Ро) саналады-триптофон опероны.
-егер ортада бұл өнім (Ро) болмаса ақуыз - репрессор оператормен қосыла алмайды, сондықтан РНҚ-полимераза оперон гендерін транскрипциялайды-оперон «іске қосылады» және Ро өнімінің түзілуі үшін қажет ферменттер ситезделінеді;
-егер ортада ақырғы өнім (Ро) (триптофан) жинақталса, оның біршама белігі ақуыз репрессормен байланысып оның оператормен қосылуына ықпал етеді- оперон «өшіріледі», Ро өнімнің түзілуіне қажет ферменттер синтезі тоқталады.
Лактоза оперонында да, индукцияланатын оперон сияқты, гендердің экспрессиясын реттеудің 2 әдісі қолданылады: 1) РНҚ-полимеразаның промотормен байланысуын реттеу (промотордың, б-фактордың, РНҚ-полимеразаның ерекшеліктері, арнайы ақуыз САР-арқылы); 2) промотормен байланысқан РНҚ-полимеразаның өз гендеріне қарай жылжуын реттеу (45-сурет).
Е. соІі жасушасында лактоза оперонының реттелуін алғаш зерттеген Жакоб және Моно (1961) болатын. Е. соІі тіршілігі үшін қалыпты энергия көзі болып глюкоза саналады. Егер де тіршілік ортасында глюкоза болмаса ол лактозаны пайдалануға көшеді. Осы кезде жасушада лактозаны ыдырататын β-галоктозидаза ферменті синтезделуі қажет. β-галактозидаза ферменті дисахарид лактозаны галактозаға және глюкозаға ыдыратады.
Ішек бактериясы (Е. соІі) жасушасында β-галактозидаза ферменттерінің синтезделуі қоректік ортада лактоза болған жағдайда индукцияланады, ал оның мөлшері азайса, не мүлдем болмаса, бұл ферменттің еинтезделу қарқыны да азаяды не тоқталады. β -галактозидаза ферменті синтезделу үшін ішек бактериясының ДНҚ,-сының Lас-Ζ гені транскрипцияланып, оның а-РНҚ-сы түзілуі қажет. β-галактозидаза ферментінің синтезделу қарқыны индукцияланғаннан кейін 1000 есеге дейін артады және ол қоректік ортада индуктор —лактоза болса бір деңгейде ұзақ уақыт ұсталып тұрады. β-галактозидаза ферментінің лактозадан басқа және негізгі индукторы ретінде оның ыдырауында пайда болатын арлық зат —аллалактоза да саналады.
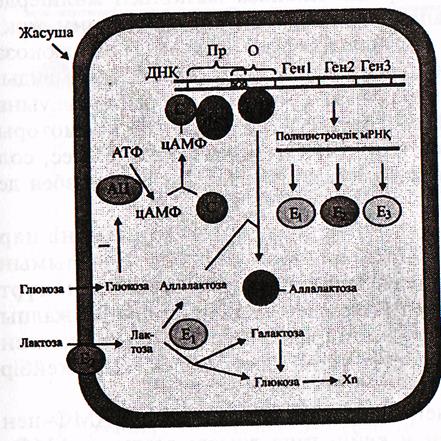
25-сурет. лактоза оперонының құрылысының жобасы (Мушкамбаров, Кузнецовтан, 2003)
Пр-промотор; О-оператор; С-САР (катаболизмді активтендіретін ақуыз); Е-РНҚ-полимераза; Rл-лактоза оперонының репрессоры; Е1—галактозидаза Е2-пермеаза, Е3-трансацетилаза; АЦ-ацетилатциклаза.
Ортада индуктордың (лактоза не аллалактоза) азаюы не жойылуы β -галактозидаза а-РНҚ-сының нуклеотидтерге ыдырап жойылуына алып келеді. Ал а-РНҚ-ның тіршілік ұзақтығы бірнеше минутқа ғана тең, сондықтан да оның бір деңгейде синтезделіп тұруы үшін, ол үнемі индукцияланып тұруы қажет, яғни ортада лактоза не аллалактоза болуы қажет.
Лактоза оперонында 3 ген болады, олардың екеуі- Lас-Ζ+ және Lас-У+лактозаны ыдырататын β-галактозидаза жене пермеаза ферменттерін кодтайды. (3-галактозидаза ферменті лактозаның галактозаға және глюкозаға ыдырауын катализдесе, пермеаза ферменті лактозаның сыртқы ортадан бактерия жасушасына енуі үшін қажет. Осы екі генмен қатар орналасқан және бірге транекрипңияланатын үшінші Lас-Ζ+ гені болады, ол транеацетилаза ферментін кодтайды, бірақ ол лактозаның ыдырауына қатынаспайды.
Лактоза оперонының негізгі реттеушісі болып лактоза емес аллалактоза саналады, себебі—ол лактоза репрессорымен байланысып оны активсіздендіреді, яғни оның операторды жауып (тығындап) тастауын болдырмайды, сондықтан да оперон гендері транскрипцияланады.
Мұның бәрі қоректік ортада глюкоза болмаған жағдайда ғана байқалады. Ал егер, қоректік ортада глюкоза жеткілікті мөлшерде болатын болса, онда лактозаны пайдаланудың еш бір қисыны жоқ, яғни биологиялық тұрғыдан алғанда тиімсіз. Сондықтан да глюкоза лактоза оперонының активтенуіне кедергі келтіреді, тіпті болдырмайды.
Бұл құбылыс РНҚ полимеразаның промотормен байланысуына әсер ету арқылы жүзеге асады. Лактоза оперонының промоторы ұзын, кең болады және ол тек қана РНҚ- полимеразамен емес, сол сияқты, ерекше ақуыз САР-катабализмді активтендіретін ақуызбен де байланысады.
Егер САР болмаса РНҚ-полимераза промотормен нашар байланысады, ал егер САР болса ол промотордың құрылымын өзгертіп оның РНҚ-полимеразамен байланысу қабілетін күрт жоғарылатады. САР эукариоттар гендеріндегі транскрипцияның жалпы факторлары сияқты рөл атқарады, тек кез-келген эукариоттар гендерінің қызмет етуі үшін қажет болеа, САР кейбір оперондар үшін ғана қажет.
САР ақуыздың промотормен байланысуы тек ЕАР+ ң-АМФ-пен қосылып кешен пайда еткеннен кейін ғана жүзеге асады. ң-АМФ-АТФ-тен аденилатциклаза ферментінің қатынасуымен түзіледі.
Қоректік ортада глюкоза болмаса аденилатциклаза ферментінің белсенділігі жоғары деңгейде болып жасушада цАМФ концентрациясы жеткілікті мөлшерде болады, сондықтан да САР лактоза промоторымен байланысады және оған РНҚ-полимераза жеп-жеңіл жалғанады. Бүл кезде оперонның белсенділігі оператордың бос болу болмауына байланысты, яғни ортада лактозаның не аллалактозаның болуына
байланысты.
Егер қоректік ортада глюкоза болса аденилатциклаза ферментінің белсенділігі төмен болып, промотор САР ақуызымен байланыса алмайды және ол РНҚ—полимеразамен де қосыла алмайды. Лактоза опероны іске қосылмай, қорек ретінде глюкоза пайдаланылады.
Триптофан оперонында да екі жақты ретгелу механизмі болады. Біріншіден РНҚ-полимеразаның оператор арқылы жылжуы реттелінеді, екіншіден (негізгісі)- транскрипцияның аттенюатор учаскесінде аяқталуы арқылы реттелінеді.
Аттенюатор - кейбір оперондарда оператор мен гендер арасында болатын ДНҚ-ның ерекше учаскесі. Бұл жерде, кейбір жағдайларда, оперон транскрипциясы аяқталады.
Аттенюаторлары бар
оперондар негізінен репрессияланатын оперондар қатарына жатады және кейбір сирек кездесетін аминқышқылдардың (триптофан, гистидин, фенилаланин) синтезделуі үшін қажет компоненттердің синтезделуін (анаболизм) қадағалайды (46-сурет).
Бұл оперондарда промотор мен оператордан кейін лидерлік бөлім деп аталатын ерекше бөлім болады, ол аттенюатормен аяқталады.
Осы бөлімнің транскрипциясы нәтиже 
 сінде лидерлік учаскенің а-РНҚ-сы түзіледі. Ол рибосомамен байланысып, трансляцияланып, лидерлік пептитті (ЛП) синтездейді. Лидерлік пептид 14 аминқышқылдары қалдықтарынан тұрады, оның екеуі триптофан-аминқышқылы. Егер жасушада триптофан аминқышқылы жеткілікті болса, лидерлік пептид (ЛП) үзіліссіз синтезделінеді, оның рибосомасы және РНҚ-полимеразасы аттенюаторға жеткен кезде транскрипцияның аяқталуы туралы сигнал пайда болады. РНҚ-полимераза ДНҚ молекуласынан диссоциацияланады (ажырайды) және ген ақпараты әрі қарай транскрипцияланбайды. Осылайша, триптофан лидерлік пептидке қосылып, аттенюаторлық механизм арқылы өзінің түзілуіне қажет ферменттердің синтезделуін бастырмалайды (репрессиялайды).
сінде лидерлік учаскенің а-РНҚ-сы түзіледі. Ол рибосомамен байланысып, трансляцияланып, лидерлік пептитті (ЛП) синтездейді. Лидерлік пептид 14 аминқышқылдары қалдықтарынан тұрады, оның екеуі триптофан-аминқышқылы. Егер жасушада триптофан аминқышқылы жеткілікті болса, лидерлік пептид (ЛП) үзіліссіз синтезделінеді, оның рибосомасы және РНҚ-полимеразасы аттенюаторға жеткен кезде транскрипцияның аяқталуы туралы сигнал пайда болады. РНҚ-полимераза ДНҚ молекуласынан диссоциацияланады (ажырайды) және ген ақпараты әрі қарай транскрипцияланбайды. Осылайша, триптофан лидерлік пептидке қосылып, аттенюаторлық механизм арқылы өзінің түзілуіне қажет ферменттердің синтезделуін бастырмалайды (репрессиялайды).
Ал егер, жасушада триптофан концентрациясы төмен (аз) болса, рибосомада лидерлік пептидтің синтезделуі кешеуілдейді және ол РНҚ-полимераза ферментіне ілесе алмай артта қалып қояды. Бұл ДНҚ-ның және а-РНҚ-ның лидерлік бөлімінің конфигураңиясын өзгертіп, аттенюаторда трансляцияның аяқталуы туралы сигналдың пайда болуын іске асырмайды. РНҚ-полимеразаның әр-бір молекуласы осы «қауіпті» учаскеден аман-есен өтіп гендерді транскрипциялайды, яғни а-РНҚ-лар синтезделінеді. Аттенюаторлық механизм триптофан оперонының белсенділігін толық бастырмаламайды (репрессияламайды)-шала бастырмалайды. Бұл оперонның толық бастырмалануы триптофан концентрациясының өте жоғары дәрежеде болған кезде ғана жүзеге асады. Бұл кезде триптофан арнайы ақуыз- репрессормен байланысып, оның оператормен қосылу мүмкіндігін жоғарылатады, триптофан+репрессор кешені оперонды тығындап, жауып, оны толық бастырмалайды (репрессиялайды), сондықтан гендер ақпараты транскрипцияланбайды.
№15 дәріс
Тақырыбы: Геном және оның құрылысы
Жоспар: 1. Прокариоттар және эукариоттар геномы
2. Адам геномы
Геном — жасушаның, ағзаның тіршілігі және дамуы үшін қажет барлық генетикалық ақпарат жазылған ДНҚ молекулаларының толық жиынтығы болып табылады, яғни жасушаның ядролық және цитоплазмалық ДНҚ-сының барлық гендері мен ген аралық учаскелерінің жиынтығы.
Геном құрылысының жалпы принңиптерін және оның құрылымдық —қызметтік ұйымдастырылуын зерттейтін ғылымды геномика деп атайды.
Адам геномикасы — молекулалық медицинаның негізі болып, тұқым қуалайтын және тұқым қуаламайтын ауруларды анықтау, емдеу және алдын-алу, болдырмау әдістерін қалыптастыру үшін маңызды рөл атқарады. Геномиканың негізгі бөлімдері: құрылымдық, қызметтік, салыстырмалы, эволюциялық және медициналық геномика.
Прокариоттар геномы — ішек бактериясында-Е.соІі, жақсы зерттелген. Бактерия хромосомасы 3,2х106 н.ж. тұратын сақиналы ұзын ДНҚ. Бактерия гендері сызықты орналасқан. ДНҚ репликациясы ТЕТА репликация типімен оrі - нүктесінен басталады. Хромосома-инициация сайтымен бірге өздігінен репликацияланатын молекула-репликон болып табылады. Бактерия геномында 2500-дей гендер болады.
Гендер белсенділігі (экпрессиясы) оперон типі сияқты реттелінеді, себебі бактериялар гендерді оперондық құрылымға ие. Ішек бактериясында (Е.соІі) бактерия хромосомасының репликонынан басқа да репликондар кездеседі, мысалы эписомалар және плазмидалар. Плазмида — бактерия хромосомасынан тәуелсіз репликацияланатын сақиналы хромосомалық элемент, оның өлшемі шамамен бактерия хромосомасының 10-20 %-дай, 1-3 гені болады.
Ең негізгі плазмидаларға бактериялардың антибиотиктер әсеріне төзімділігін қалыптастыратын төзімділікті тудырушы факторлар жатады, олар 10-15 көшірме күйінде кездеседі. Эписомалар—бактерия хромосомасынан бөлек, автономды кездесетін не оған жалғанатын сақиналы хромосомалық элементтер. Ең жақсы зерттелген эписома, бұл Ғ-фактор (фертильдік фактор). Ол бактериялардың жыныстық процесін анықтайды және аталық жасушаларда (Ғ+жасушалар) кездеседі. Эписомалардың кейбіреулері инфекциялы болып келеді. Егер эписомаларда антибиотиктерге төзімділікті қалыптастыратын гендер болса, онда олар бактерия жасушаларына жеп-жеңіл өтіп, медицина үшін үлкен проблемалар туғызады. Бактерия геномында қозғалғыш генетикалық элементтерде кездеседі.
Эукариоттар геномы — көлемі және құрылысы жағынан күрделі болады, олар; нуклеотидтер—кодондар—гендер мен ген аралық учаскелер—күрделі гендер—хромосома иіндері—хромосомалар—гаплоидты хромосома саны сияқты бірте-бірте күрделенетін құрылымдардан тұрады. Эукариоттар геномының көлемі өте үлкен болады, себебі олардың нуклеотидтер бірізділігі (ДНҚ молекуласы) тек қана қайталанбайтын учаскелер емес, сол сияқты орташа қайталанатын және өте жиі қайталанатын учаскелерден тұрады. Сол сияқты, геномның өте үлкен болуын гендердің экзон-интрондық құрылысымен де түсіндіруге болады. Эукариоттар геномы ядролық және ядродан тыс орналасқан ДНҚ молекулаларынан тұрады. Соңғысына цитоплазманың сақиналы ДНҚ-сы: плазмидалар, эписомалар, митохондрия және пластидтер ДНҚ-сы жатады. Ядролық ДНҚ хромосомасынан тыс орналасқан гендер жиынтығын плазмондар деп атайды, олар цитоплазмалық тұқым қуалаушылықты анықтайды. Ядролық ДНҚ-массасының бәрі дерлік хромосомаларға таралған. Хромосомалар құрылысы күрделі.
Эукариоттар геномына қозғалғыш генетикалық элементтер — транспозондар да тән, олар гендер белсенділігін реттеуге қатынасады, яғни бұрын пассив күйде болып келген гендерді активтендіреді немесе керісінше.
Адамның сома жаеушасындағы (2n) ДНҚ-ның жалпы мөлшері 6,4.109 н.ж. тең, яғни гаплоидтық хромосома жиынтығында (n)-3,2.109 н.ж. ДНҚ молекуласының 99,5 хромосомаларда кездеседі және бұл ядро ДНҚ-сы болып табылады. Ядродан тыс ДНҚ молекуласы-митохондрияларда, цитоплазмада (0,5)-сақиналы ДНҚ күйінде кездеседі.
ХХ-ғасырдың 60-жылдары Р.Бриттен және Э.Дэвидсон эукариоттар геномының молекулалық құрылысының ерекшеліктерін, яғни геномның әртүрлі учаскелерінің түрліше рет қайталанатынын ашты. ДНҚ молекуласының қайталанбайтын, орташа қанталанатын, өте жиі қайталанатын учаскелері белгілі. Кайталанбайтын учаске ДНҚ молекуласының бойында бір дана күйінде кездеседі және бүл жерлерде барлық етруктуралық гендер орналасқан. Оның үлесіне ДНҚ молекуласының 75 көлемі тиесілі. Геномның қалған 25% - қайталанатын нуклеотидтер бірізділігі болып табылады. Олар жүзден мыңдаған ретке дейін қайталануы мүмкін. Оларды дисперсияланған (біркелкі таралған) және сателиттік ДНҚ бірізділіктері деп бөледі.
Дисперсияланған (біркелкі таралған) ДНҚ бірізділіктері (геномның 15% көлемін құрайды) ДНҚ молекуласының бойына біркелкі бытыраңқы таралып орналасқан. Оларға SINE (қысқа элементтер), LINE (ұзын элементтер) және басқа да бірізділіктер кіреді.
Жеке SINE- бірізділіктерінің ұзындығы 90-500 н.ж., ал LINE -бірізділіктерінің ұзындығы 7000 н.ж. дейін жетеді.
SINE - бірізділіктерінің кейбіреулерін ALu- бірізділіктері деп атайды, себебі олар ALu - рестриктазалар арқылы кесіледі. Адам геномында 300 000 нан 500 000-ға дейін АІu - бірізділіктер табылған. Бұл бірізділіктердің бір ерекшеліктері — олар өздігінен көшірмеленіп, ДНҚ-ның кез-келген бөліміне, сол сияқты гендерге, қыстырылып қосылуы мүмкін. Соңғы жағдайларда олар мутация пайда етіп ген қызметін бұзады.
Сателиттік қайталанулар хромосомалардың әр түрлі учаскелерінде бумаланып жинақталған және көптеген рет қайталанатын тандемді бірізділіктерден тұрады. Сателиттік ДНҚ геномның шамамен 10% қамтиды және а-сателиттік, минисателиттік және микросателиттік ДНҚ-лар деп бөлінеді.
α-Сателиттік ДНҚ, әдетте, барлық хромосомалардың центромераларының айналасында орналасқан. Олардың негізі 171 нуклеотидтер жұптарынан тұрады және жұптасып (тандемді) мындаған рет қайталанады.
Минисателлиттік ДНҚ - 20-70 нж. тұратын және ондаған рет жұптасып (тандемді) қайталанатын бірізділіктер.
Микросателликтік ДНҚ - 2-4 нж. тұратын, жалпы ұзындығы жүздеген нуклеотидтер жұптарынан аспайтын, жұптасып (тандемді) байланысқан қайталанулар типі болып табылады.
«Адам геномы» атты ғылыми бағдарлама ХХ-ғасырдың 90-жылдары басталып 2001-2003-жылдары толық аяқталды. Бұл бағдарламаны орындауға Қытай, Жапония, Франция, АҚШ, Ұлыбритания елдерінен 20-ға жуық ғылыми зерттеу мекемелері ат салысты. Бұл бағдарламаның негізгі мақсаты адам геномын зерттеп секвендеу (секвендеу-барлық хромосомалардағы ДНҚ молекуласының нуклеотидтер бірізділігін анықтау) және адам хромосомаларының физикалық және генетикалық картасын құрастыру болып табылады. Адам геномын секвендеу, адам геномының табиғи нұсқаларын талдау, ең жиі кездесетін полиморфизм-жекелеген нуклеотидтер полиморфизімін, ашуға, полиморфизм картасын құрастыруға мүмкіндік берді. Жекелеген нуклеотидтер полиморфизмі дегеніміз-ДНҚ молекуласының бір бөлімінде бір нуклеотидтің екінші бір нуклеотидпен алмастырылуы. Бұл фермент белсенділігінің өзгеруіне алып келеді. ЖНП-ДНҚ-ның әрбір килобазасында (1 кб=1000 нуклеотидке тең) кездеседі. Адам геномының ұзындығы 3,2 млрд н.ж. тең десек, онда геномда кездесетін ЖНП халпы саны 1,6-3,2 миллиондай болады. Олардың 2,5 миллионға жуығы анықталды. Әрбір адам бір-бірінен гем құрамында кездесетін бір нуклеотидтер жұбының өзгеше болуы арқылы ерекшелінеді және бұл адамдар фенотипінің сан алуан түрлі болуына алып келеді. ЖНП қартасын құрастыру мультифакторлы полигенді патологиялардың, мыс. рак, диабет, психикалық аурулар т.б. дамуына жауапты гендерді идентификациялауға мүмкіндік берді.
Қазіргі таңда адамның 3000-нан астам тұқым қуалайтын ауруларының нақтылы гендерінің орналасқан жерлері анықталды, 20 мыңдай гендердің хромосомаларда орналасу орны белгілі болды, көптеген хромосомалық делециялық синдромдардың себептері анықталды. ЖНП-нің көпшілігі гендер экзондарында кездеседі.
Адам геномын зерттеулер нәтижесінде қазіргі таңда біз өз гендеріміздің 50% -ының құрылысын, қызметтерін жақсы білеміз, қалғандары белсенді түрде зерттелуде және жақын арада анықталады деп күтілуде. Бүгінгі күні кез-келген адам өзінің генетикалық төлқұжатын жасатып, соған сәйкес салауатты өмір сүру бағдарламасын құрастыруға мүмкіндік алып отыр. 2000-2003 жылдан бері қарай адамзат постгеномдық дәуірде тіршілік етуде, себебі осы жылы «адам геномы» атты халықаралық ғылыми бағдарлама табысты аяқталды (Ф.Коллинз, 2000). Бұл бағдарламаның аяқталуы генетиканың әрі қарай дамуының 3 жаңа стратегиясын қалыптастырды: 1) генетика -медицина үшін (пренатальдық диагностика, тұқым қуалайтын аурулар);
2) генетика —денсаулық үшін (аурулардың алдын алу -болдырмау);
3) генетика қоғам үшін (дәрігерлерге, көпшілікке генетиканы үйрету).
Жоғарыда айтылғандардың бәрі ядро хромосомаларындағы геномға
жатады. Сонымен қатар, адам геномы митохондрия геномын және цитоплазмада, ядрода кездесетін сақиналы ДНҚ молекулаларын да камтиды.
Митохондрия ДНҚ-сының (мтДНҚ) геномы 165 н.ж. тұратын қос тізбекті сақиналы молекула болып табылады. Әрбір митохондрияда 10-шақты ДНҚ молекуласы кездеседі. мт-ДНҚ-сында интрондар болмайды, оның ұүрамында 2р-РНҚ, 22-т-РНҚ және 13 фосфорлау полипептидтерінің гендері кездеседі. Митохондрий геномы 1981 ж. толық анықталған (47-еурет). Адамның сақиналы ДНҚ-сы толық зерттелмеген оның өлшемі 150 н.ж.-тан -20000 н.ж. дейін болады. Ядроның сақиналы ДНҚ-сы онкогендермен уларға төзімділік гендерінің амплификацияланған (көшірме-ленген) учаскелері болып табылады. Адам геномының жалпы ұзындығы 3000-3500 см тең.
13. МОЛЕКУЛАЛЫҚ БИОЛОГИЯ ПӘНІНЕН ТЕСТ СҰРАҚТАРЫ.
1. Жасушаның митоздық боліну құбылысындағы реті:
телофаза
интерфаза
метафаза
анафаза
профаза
2. Жасушаның даму кезендерінің сатылары:
цитокинез
интерфаза
митоз
Нуклеин қышқьшдардың мономерлері
Құрамында 7 көміртегісі бар моноқанттар деп аталады
Рибосомалардың қүрамына кіретін заттар:
ақуыздар
липидтер
ДНҚ
4. РНҚ
6. Цитоплазманың қозғалысы:
циклоз
таксис
тропизм
7. Тұқым куалау ақпаратын тасымалдаушы:
ДНҚ
РНҚ
АТФ
8. Микроскоптың үлкейтетін болігі:
штатив
тубус
окуляр
револвер
9. Центромерлердің бөліну ұршық экваторда орналасқан митоз
кезеңі:
интерфаза
профаза
метафаза
анафаза
телофаза
10. Жасушадағы ақуыздардың орташа мөлшсрі:
80 %
20 %
1 %
11. Полисомалардың түзілетін, жіктелетін орны:
цитоплазмада
ядрода
3. вакуольде
12. Акуыздың биосинтезі кезінде рибосомада түзілетін құрылым:
полипептидті құрылым
екінші реттік құрылым
үшінші реттік құрылым
13. Хромосомалар центромерлермен екі полюскс тартыла бастаған
митоз кезеңі:
интерфаза
профаза
метафаза
анафаза
телофаза
14. Генетикалық ақпаратты бір ұрпақтан екінші ұрпаққа
тасымалдаушы:
акуыздар
рибосомалар
хромосомалар
15. Ақуыздардың күрделі құрылыстары түзелетін орын:
хлоропластарда
рибосомаларда
митохондрияларда
эндоплазмалық торда
16. Рибосомалардың түзілетін, жиналатын орны:
ядро шырынында
ядрошықга
3. ядроның жарғақшаларында
17. Ақуыз молекуласының күрделі құрылымдарға түзейтін орны:
рибосомаларда
цитоплазманың матриксінде
эндоплазмалық тордың өзекшелерінде
18. Центриольдерде бөліну ұршығы пайда болған митоз ксзеңі:
интерфаза
профаза
метофаза
анафаза
телофаза
19. Генетикалық кодтың үшөрімді екенін дәлелдеп, 1961 жылы
ұсынған ғалым:
Ф.Крик
Д.Уотсон
Э. Чаргафф
Л.Полинг
М.Уилкинс
20. Ақуыз молекуласының бірінші құрылысындағы
аминқышқылдарының байланысы:
дисульфидті байланыста
пептидті байланыста
сутегілік байланыста
21. Рибосомалардың синтезделетін қосылыстар:
акуыз
майлар
АТФ
4. көмірсулар
22. Митохондриядағы ферменттердің, ДНҚ, РНҚ және рибосомалардың болатын орны:
кристаларда
матриксте 3. сыртқы жарғақшасында
23. Полиқанттардың мономерлері - @@ деп аталады
24. Құрамында 6 көміртегісі бар моноқаннтер @@ деп аталады
Гликоген жоне целлюлоза @@ полимерлері
ДНК молекуласы екі @@ тізбектен тұрады
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 3848; Нарушение авторских прав?; Мы поможем в написании вашей работы!