КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Метаболизм в мышечной ткани
|
|
|
|
Химизм мышечного сокращения
Обмен веществ покоящихся мышц незначителен, при работе он повышается более чем в 100 раз. Для сокращения мышц необходимы: 1)гликоген, 2) миозин, 3) актин, 4) АТФ, 5) креатинфосфат, 6) ионы калия.
Химизм мышечного сокращения в настоящее время рассматривается как результат взаимодействия между собой двух сократительных белков - актина и миозина за счет расщепления АТФ АТФ-азой миозина. Энергия, выделяющаяся при расщеплении АТФ, превращается из химической в механическую, нити актина вытягиваются между нитями миозина, саркомер укорачивается или напрягается. В условиях покоя АТФ не расщепляется, так как АТФ-азная активность миозина подавлена присутствием ионов магния. Роль магния – тормозить в условиях покоя гидролиз АТФ, при этом головки миозина получают энергию, которая обеспечивает смещение тропонина С и последующее зацепление ее за актиновую нить.
Стимулом для включения сложного механизма служит нервный импульс, при его поступлении в синапс (на окончание двигательного нерва), выделяется медиатор - ацетилхолин – это создает разность потенциалов между наружной и внутренней мембраной. Вследствие этого выходит кальций из саркоплазматического ретикулума, где он находится в связи с белком кальсеквестрином, что обеспечивает увеличение его содержания в саркоплазме. Кальций действует, как антагонист магния, создает условие для проявления АТФ-азной активности миозина. АТФ расщепляется, сократительные белки взаимодействуют, мышца (саркомер) сокращается. Сокращение продолжается до тех пор, пока не поступают нервные импульсы и есть запасы АТФ. За счет энергии АТФ ионы кальция связываются саркоплазматическим ретикулумом, удаляется из саркоплазмы, их концентрация снижается. Происходит расслабление мышц. Поддержанию АТФ в должной концентрации обеспечивает высокий уровень содержания дыхательных ферментов. Увеличение кровоснабжения мышц при работе, ведет к увеличению притока кислорода, и питательных веществ. В начальный период может быть использован кислород, миоглобин, креатинфосфат, затем глюкоза, гликоген, липиды. Это обеспечивается высокой концентрацией гликогена и активностью ферментов гликолиза.
Обмен веществ в состоянии покоя находится на относительно низком уровне, так как мышцы функционируют непостоянно. Метаболизм работающей мышцы направлен только на обеспечение мышечной работы и его специфика в первую очередь состоит в производстве молочной кислоты из гликогена, содержание которого достигает 1% от общего веса мышц. Здесь идет захват глюкозы, регулируемый инсулином и задерживаемый клеткой в виде глюкозо-6-фосфата гексокиназой с Кm = 10-5моль/л. Таким образом этот фермент фосфорилирует глюкозу до тех пор, пока глюкозо-6-фосфат не вступит во внутренний цикл: гл-6-ф↔гликоген. Известно, что гликоген распадается при увеличении концентрации глюкагона и адреналина в крови, однако сарколемма не имеет рецепторов для глюкагона и гликоген распадается при действии адреналина, причем аденилатциклаза обладает более высокой чувствительностью, нежели в печени. При расщеплении количества гликогена, находящегося в 1г мышцы до лактата освобождается 1-2 ккал энергии могущей быть использованной в сокращении мышцы. Интенсивная мышечная работа ведет к образованию большого количества молочной кислоты, которое может достигнуть 100 г.
Накопление лактата ограничивает гликогенолиз и для ликвидации молочной кислоты требуется значительный период повышенного обмена. Происходит переход на аэробное окисление, благодаря увеличению транспорта кислорода в работающую мышцу и лактат окисляется до пирувата, который даст глюкозу и затем гликоген (в период отдыха), или в сердечной мышце окисляется до СО2 и Н2О при ликвидации “кислородной задолженности” с выделением большого количества энергии в цикле Кребса. Таким образом, молочно-кислое брожение идет в саркоплазме, а терминальное окисление – в митохондриях. В мышечной ткани особенно активны изоферменты лактатдегидрогеназы, причем ЛДГ1 сдвигает равновесие между лактатом и пируватом в сторону пирувата (в сердечной мышце) – начального метаболита аэробной стадии, а ЛДГ5 – в сторону лактата – конечного продукта анаэробного гликолиза (в скелетной мускулатуре).
Дефекты в структуре ферментов, расщепляющих гликоген, приводят к гликогенозам. Так, в результате отсутствия или дефекта α1-4 гликозидазы (болезнь Помпе) вызывает слабость скелетных мышц, расширение сердца и смерть в раннем возрасте. Наиболее из гликогенозов распространена болезнь Мак-Ардля – следствие дефекта мышечной фосфорилазы, приводящего к судорогам при тяжелой мышечной работе. Симптомы выражены незначительно, так как мышца переходит на потребление энергии β-окисления жирных кислот при интенсификации липолиза. Т.к. β-окисление идет в митохонжриях, для транспорта жирных кислот из саркоплазмы необходим карнитин, концентрация которого в мышцах очень велика.
β - окисление жирных кислот очень выгодный аэробный процесс, который доминирует при работе красных волокон сердечной мускулатуры. Здесь же хорошо сгорают кетоновые тела, сохраняя чистую глюкозу для питания мозга, куда не поступают жирные кислоты через гематоэнцефалический барьер. Катехоламины ускоряют липолиз и β-окисление, в результате увеличения содержания жирных кислот в крови. Кетоновые тела, образующиеся при неполном окислении жирных кислот в печени, в мышцах (особенно сердечной) тоже являются источником энергии. Для сердечной мышцы характерна реакция превращения ацетоацетата в энергетический продукт:
митохондриальный фермент внепеченочных тканей
 CH3-CO-CH2-COOH + НOOС-СН2-СH2-CO~SkoA
CH3-CO-CH2-COOH + НOOС-СН2-СH2-CO~SkoA
CH3-CO-CH2-СO~SkoA + HOOC-CH2-CH2-COOH
ацетоацетил koA сукцинат
НSkoA
CH3-CO-CH2-СO~SkoA → 2СН3СО~SkoA→(АТФ) в ЦТК
тиолаза
Схема химизма мышечного сокращения
ЦНС - - - → возбуждение - - → синапс
↓
 ацетилхолин
ацетилхолин
потенциал действия
выделение ионов кальция из цистерн саркоплазматического ретикулума
↓
 взаимодействие кальция с Тn(с) переход его во включенное состояние
взаимодействие кальция с Тn(с) переход его во включенное состояние
зацепление головок миозина за актин → актомиозин
SR - Ca++ 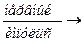 SR + Ca++ - Выход Са++ из саркоплазматического
SR + Ca++ - Выход Са++ из саркоплазматического
ретикулума.
A – T + Ca++ 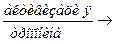 A + T – Ca++ - активация актина
A + T – Ca++ - активация актина
А + М – АТФ 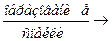 М – АТФ – А – образование актомиозина
М – АТФ – А – образование актомиозина
М – АТФ – А + Н2О 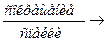 М – АДФ – А + Фн + работа – изменение угла соединения головки миозина с 90˚ до 45˚
М – АДФ – А + Фн + работа – изменение угла соединения головки миозина с 90˚ до 45˚
М – АДФ – А + АТФ 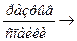 М – АТФ + А + АДФ
М – АТФ + А + АДФ
М – АТФ имеет низкое сродство к А
Т – Cа++ + SR + АТФ 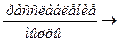 Т + SR – Ca++ + АДФ + Фн – связывание Са++
Т + SR – Ca++ + АДФ + Фн – связывание Са++
снова с саркоплазматическим ретикулумом.
Источники энергии для работы мышц. Некоторый резерв энергии находится в небольших количествах АТФ и креатинфосфата, который есть в мышце. Этого запаса хватает на 10-12 с. Дополнительное количество АТФ образуется под действием аденилаткиназы, которая катализирует реакцию: 2АДФ→АТФ+АМФ
АМФ в свою очередь является аллостерическим фактором фосфофруктокиназы - повышающим скорость гликолиза в целом, таким образом к 20-й секунде активируется гликогенолиз, который за счет субстратного фосфорилирования обеспечивает работающую мышцу молекулами АТФ и достигает максимума через 40-50 секунд непрерывной работы мышц. Через 60-70 секунд доминируют уже аэробные процессы, благодаря увеличению транспорта кислорода в работающую мышцу. При аэробном окислении образуется АТФ в митохондриях, которые в большом количестве окружают мышечное волокно. АТФ поставляют в основном ЦТК и β-окисление жирных кислот.
Источники энергии для работы мышцы
|
|
|
|
|
Дата добавления: 2015-07-02; Просмотров: 2453; Нарушение авторских прав?; Мы поможем в написании вашей работы!