КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Антиоксидантная защитная система растений
|
|
|
|
Согласно литературным данным, в растительной клетке существует 2 механизма обезвреживания АФК [38]:
1. Ферментативный, представленный работой ферментных систем, обезвреживающих АФК – супероксиддисмутаза (СОД), каталаза, пероксидаза.
2. Неферментативный, включающий низкомолекулярные антиоксиданты, такие как глутатион, аскорбат, пролин, каротиноиды, токоферол и др.
СОД катализирует диспропорционирование супероксидных-анион радикалов до молекулярного кислорода и пероксида водорода. В клетках растений существуют 3 основные изоформы СОД:
Cu-Zn-СОД – гомодимер (33кДа) в активном центре по 2 атома меди и цинка. Локализация - хлоропласты, митохондрии, пероксисомы, цитоплазма, апопласт.
Mn-СОД – гомодимер (46кДа) или гомотетрамер (92кДа) в активном центре содержится 2 или 4 атома марганца. Локализация - митохондрии, пероксисомы.
Fe-СОД – гомодимер (36кДа-46кДа) – в хлоропластах. Содержание варьирует от 1 до 2 атомов железа в активном центре [64].
Механизм действия всех изоформ СОД заключается в последовательном восстановлении и окислении супероксидными-анион радикалами металла активного центра фермента. Поскольку гидроксильные радикалы, синглетный кислород активно окисляют белковые молекулы, специфических ферментов-дезактиваторов данных АФК не существует, уровень их в клетке опосредованно регулируется СОД путем уборки супероксидных радикалов – источника их образования. По этой причине СОД рассматривается учеными как первичная линия защиты от окислительных повреждений, которая обрывает окисление клеточных макромолекул еще на стадии инициирования [3].
Каталаза представляет собой гемовый фермент, состоящий из четырех субъединиц с общей молекулярной массой около 240 кД. Это металлопротеин с гемовым железом в активном центре. В растениях обнаружено 5 изоформ каталаз. Необходимы достаточно высокие концентрации Н2О2 для проявления каталитической функции фермента, имеющего невысокое сродство к субстрату и использующего в качестве донора электрона при восстановлении Н2О2 до воды и молекулярного кислорода вторую молекулу пероксида. Согласно литературным данным, каталаза может разложить 44000 молекул Н2О2 в секунду [32].
Пероксидазы различаются по субстратам окисления и строению активного центра. Это металлопротеин, содержащий железо в гемовой форме. Пероксидаза катализирует окисление перекисью водорода или молекулярным кислородом ароматических аминов, анилина, бензидина, тирозина, триптофана, индола, индолилуксусной кислоты, фенолов и т.д. Источником АФК для работы пероксидазы могут быть также органические гидроперекиси. Наиболее распространена в растениях гваяколовая пероксидаза, участвующая в биосинтезе лигнина. Особое место занимает глутатионпероксидаза, использующая водород сульфгидрильной группы глутатиона для восстановления гидроперекисей липидов в соответствующие спирты и восстановление пероксида водорода до воды [10]. Структурно ферменты данного семейства представляют собой селеносодержащие тетрамеры. В активном центре фермента находится остаток аминокислоты селеноцистеина.
Неферментативным антиоксидантным звеном является глутатион, который представляет собой небольшой пептид, состоящий из трех аминокислотных остатков (-Глу, -Цис, -Гли), гамма-глутамилцистеинилглицин. Необычный структурный элемент молекулы – амидная связь, образованная взаимодействием амидной группы цистеина и карбоксильной группы глутаминовой кислоты. Глутатион защищает тиольные группы белков, инактивирует радикальные частицы, разрушает перекисные соединения, реагирует с активными формами кислорода [12]. Он может стабилизировать мембранные структуры, удаляя ацильные перекиси, образующиеся в ходе ПОЛ. При этом восстановленная форма глутатиона GSH превращается в окисленную GS-SG. Восстанавливается окисленный глутатион под действием фермента ГР. Глутатион является кофактором ферментов ГП – восстанавливает H2O2 до воды при окислении глутатиона. Снижение количества восстановленного глутатиона может наблюдаться в результате недостатка глутатионредуктазы и/или снижения скорости синтеза. Это может приводить к сильным клеточным повреждениям. Уменьшение пула восстановленного глутатиона может свидетельствовать о его окислении до GS-SG катализируемом тяжелыми металлами, либо использовании GSH-пероксидазами в процессе ферментативной регенерации перекисей липидов [4].
Витамин С (аскорбиновая кислота) также является антиоксидантом неферментативного звена и участвует с помощью двух различных механизмов в ингибировании ПОЛ. Во-первых, аскорбат восстанавливает окисленную форму витамина Е и таким образом поддерживает необходимую концентрацию этого антиоксиданта непосредственно в мембранах клеток. Во-вторых, он, будучи водорастворимым витамином и сильным восстановителем, взаимодействует с водорастворимыми активными формами кислорода НО•, О2•-, Н2О2, и инактивирует их при этом окисляясь до кислоты МДГА или ДГА [31]. Окисленные формы снова превращаются в востановленный аскорбат с помощью редуктаз МДГА и ДГА соответственно. В первом случае используется восстановленный эквивалент НАДФН+H+, а во втором – глутатион.
Аскорбиновая кислота найдена в хлоропластах, цитозоле, вакуолях и внеклеточных компартментах клетки. По литературным данным, в хлоропластах мезофилла листа находится около 20-40% аскорбата [31]. Там же содержатся все ферменты регенерации восстановленной аскорбиновой кислоты из окисленных форм. Было выявлено, что H2O2 обезвреживается в хлоропластах в аскорбат-глутатионовом цикле (Холливела–Асады) при участии аскобатпероксидазы [1,37]. По мнению ученых, аскорбатпероксидазная реакция – это центральный процесс целого цикла реакций, направленного на удаление основных активных форм кислорода в хлоропластах.
Цикл включает в себя следующие ферменты: АП, МДАсР, ДАсР и ГР. Во время работы цикла происходит восстановление H2O2 в присутствии аскорбиновой кислоты, которая восстанавливается при окислении глутатиона. Окисленный глутатион восстанавливается при участии НАДФН [19].
Аскорбат-глутатионовый цикл подытоживает целый ряд реакций, которые направлены на фотовосстановление молекулярного кислорода до воды в ФС I электронами, образованными в ФС II из воды при ее фотоокислении.
Молекулярный механизм аскорбат-глутатионового цикла включает следующие стадии:
1. Окисление воды в ФС II:
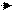 2Н2О 4е- +О2+4Н+
2Н2О 4е- +О2+4Н+
2. Фотовосстановление молекулярного кислорода до супероксидного анион-радикала в ФС I при псевдоциклическом потоке электронов:
 2e- + 2O2 2O2-•
2e- + 2O2 2O2-•
3. Реакция дисмутации супероксида до пероксида водорода и молекулярного кислорода, катализируемая СОД:
 2O2-• + 2 H+ O2+H2O2
2O2-• + 2 H+ O2+H2O2
4. Восстановление пероксида водорода аскорбатом до воды, катализируемое АП:
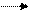 H2O2+2АсК 2Н2О + 2МДАсК
H2O2+2АсК 2Н2О + 2МДАсК
5. Регенерация окисленной формы аскорбата, МДАсК и/или ДАсК, при работе соответсвующих редуктаз МДАсР и ДАсР за счет восстановленного ферредоксина или НАД (Ф) Н из ФС I, которые в свою очередь восстанавливаются электронами из ФС II:
 2е- + 2H+ 2ДАсК АсК
2е- + 2H+ 2ДАсК АсК
Таким образом, в аскорбат-глутатионовый цикл 2 из 4 электронов от фотовосстановления двух молекул воды идут на одновалентное восстановление О2, а остальные 2 электрона используются при восстановлении окисленного аскорбата [25].
Восстановление дегидроаскорбиновой кислоты происходит с использованием протонов трипептида глутатиона. Окисленный глутатион регенерируется при работе ГР в реакции:
 GS-SG + ГР-НАДФН 2GSH+ГР-НАДФ+
GS-SG + ГР-НАДФН 2GSH+ГР-НАДФ+
При восстановлении глутатиона донором электронов служит опять фотосинтетическая НАДФН, образующийся в световых реакциях.
В аскорбат-глутатионовый цикле скорость псевдоциклического потока электронов на молекулярный кислород в ФС1 на несколько порядков ниже, чем протекание реакций дисмутации супероксида, катализируемой СОД, восстановления пероксида водорода до воды, катализируемой АП, и восстановления окисленных аскорбата и глутатиона. Поэтому аскорбат-глутатионовый цикл эффективно уменьшает время жизни фотоиндуцированных АФК – супероксида и пероксида водорода – что позволяет снизить вероятность образования гидроксильных радикалов и их взаимодействие с важнейшими биополимерами хлоропластов, и как следствие – избежать фотоингибирования [25].
Пролин – гетероциклическая аминокислота. У многих растений она накапливается в условиях биотического и абиотического стрессового воздействия. При этом пролин оказывает стабилизирующее действие на мембраны, уменьшает осмотический стресс, защищает белки от денатурации, участвует в передаче стрессового сигнала, регулирует редокс-потенциал клетки. Есть также данные, что он участвует в инактивации свободных радикалов, образуя с ними долгоживущие конъюгаты.
Еще одним важнейшим неферментативным антиоксидантным звеном растений являются каротиноиды. По химической природе каротиноиды относятся к классу тетратерпеноидов. Каротиноиды, благодаря наличию сопряженных двойных связей, могут связывать синглетный кислород и ингибируют образование свободных радикалов, предупреждая их негативное действие на растительный организм. Они обеспечивают защиту от ультрафиолетового излучения, так как могут трансформировать энергию УФ - света в видимый свет, что проявляется в явлении флуоресценции. Выступают в роли антиоксидантов, защищая чувствительные ткани и лабильные соединения от окисления [15,31].
Каротиноиды снимают избыток энергии у возбужденного хлорофилла или синглетного кислорода и рассеивают ее через процесс нерадиационной релаксации в виде тепла. Таким образом, может быть предотвращен окислительный стресс. Они так же каротиноиды могут переносить электроны с радикала и модифицировать его структуру, предотвращая дальнейшее развитие цепной реакции [57,31].
Токоферолы (витамин Е) – важнейшие природные антиоксиданты неферментативного звена. По химической природе токоферолы, как и каротиноиды, относятся к семейству терпеноидов (содержат в своей структуре изопрен). В связи с наличием в молекуле токоферолов длинной боковой изопреновой цепи все они нерастворимы в воде, но хорошо растворяются в жирах и органических растворителях [35]. Токоферолы синтезируются только в растительных клетках. Существует несколько изомерных форм токоферолов, но в наибольших количествах содержится α-токоферол. Антирадикальная активность α-ТФ обусловлена наличием в его структуре гидроксильной группы –ОН, присоединенной к ароматическому ядру и экранированной двумя метильными группами. Благодаря наличию в структуре ароматического кольца обобщенной системы π-электронов происходит смещение отрицательного заряда на кислород, результатом чего становится возможным достаточно легкий отрыв атома водорода в гидроксильной группе -ОН с образованием радикальной формы –О* альфа-токоферола. Ослаблению связей водорода с кислородом способствуют также соседние метильные группы, экранирующие гидроксильную группу. Витамин Е отдаёт атом водорода свободному радикалу пероксида липида, восстанавливая его до гидропероксида и таким образом останавливает развитие ПОЛ.
Таким образом, анализ литературных данных показывает, что в растениях существует множество систем, обеспечивающих регуляцию уровня активных форм кислорода в растительной клетке в меняющихся условиях среды. По механизму действия антиоксиданты можно разделить на несколько групп [20]:
1. "мусорщиков", которые очищают растительный организм от всех свободных радикалов, чаще всего восстанавливая их до стабильных неактивных продуктов.
2. "ловушки"- антиоксиданты, которые имеют сродство к какому-то определенному свободнорадикальному продукту (ловушки синглетного кислорода, гидроксил-радикала и т.д.). Ловушки часто используют для уточнения механизма свободнорадикальной реакции.
3.антиоксиданты, обрывающие цепи - вещества, молекулы которых более реакционноспособны, чем их радикалы. Чаще всего это фенолы, которые легко отдают свои электроны, превращая радикал, с которым они прореагировали, в молекулярный продукт, а сами при этом превращаются в слабый феноксил-радикал, который уже не способен участвовать в продолжении цепной реакции.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2523; Нарушение авторских прав?; Мы поможем в написании вашей работы!