КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Присоединение к лизосомному ферменту нескольких групп маннозо-6-фосфата усиливает сигнал сортировки
|
|
|
|
8-37
8.8.4. Лизосомные ферменты сортируются в аппарате Гольджи мембраносвязанным белком-рецептором, узнающим маннозо-6-фосфат [56]
Для образования лизосом необходим синтез специфических лизосомных гидролаз и мембранных белков. И те, и другие белки синтезируются в ЭР и транспортируются через аппарат Гольджи. Транспортные пузырьки, доставляющие их в эндолизосомы, а затем в лизосомы, отделяются от транс-сети Гольджи. Эти пузырьки должны включать именно лизосомные белки и не включать множество других белков, которые упаковываются в другие транспортные пузырьки и доставляются в другие органеллы.
Каков механизм узнавания лизосомных белков? Что обеспечивает точность отбора? Собственно, эти же вопросы можно задать и в других случаях внутриклеточной сортировки, происходящей с участием транспортных пузырьков. На молекулярном уровне ответ известен только для одного класса ферментов-лизосомных гидролаз. Они имеют уникальный маркер- маннозо-6-фосфат. который присоединяется к N-связанным олигосахаридам этих растворимых лизосомных ферментов. Реакция протекает в пространстве цис-компартмента Гольджи. Соответствующие маннозофосфатные рецепторы группируются на мембране и затем концентрируются в покрытых клатрином окаймленных пузырьках. Они также были выделены и охарактеризованы. Оказалось, что эти рецепторы представляют собой трансмембранные белки, которые связывают лизосомные ферменты, отделяя их таким образом от всех остальных белков и собирая в окаймленные транспортные пузырьки. Эти пузырьки быстро теряют свою кайму и сливаются с эндолизосомами.
В некоторых клетках небольшое количество рецепторов маннозо-6-фосфата присутствует в плазматической мембране, где они участвуют в эндоцитозе лизосомных ферментов, которые были выделены во внеклеточную среду. Благодаря этим рецепторам ферменты через окаймленные ямки попадают к эндосомам, а оттуда к лизосомам. Таким необычным путем, с помощью «старьевщиков» и доставляются в лизосомы гидролазы, которые избежали процесса упаковки в транс-сети Гольджи и были поэтому транспортированы к клеточной поверхности и выведены наружу.
8.8.5. Рецептор маннозо-6-фосфата «курсирует» между специализированными мембранами [57]
Маннозофосфатный рецептор был очищен и охарактеризован в экспериментах in vitro. Оказалось, что он связывает специфический олигосаха-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
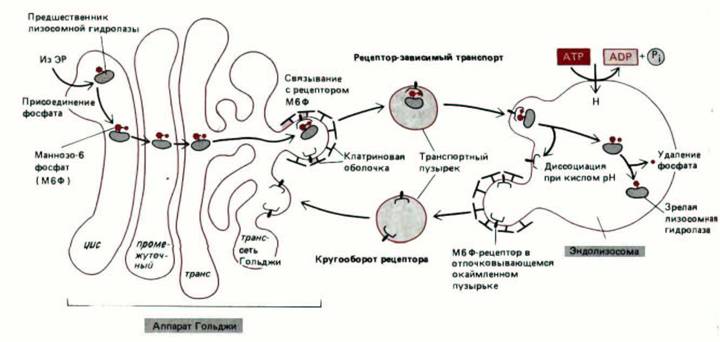
Рис. 8-72. Транспорт вновь образованных лизосомных гидролаз в лизосомы. В цис-аппарате Гольджи предшественники лизосомных гидролаз «метятся» при помощи ман-нозо-6-фосфатных групп, а в транссети Гольджи отделяются от других типов белков. Это отделение происходит потому, что отпочковывающиеся от транс-сети Гольджи клатриновые окаймленные пузырьки содержат рецепторы маннозо-6-фосфата, связывающие лизосомные гидролазы. Пузырьки утрачивают «кайму» и сливаются с эндолизосомами (см. рис. 8-71). При низком рН, который существует в эндолизосомах, гидролазы отщепляются от рецепторов. Рецепторы возвращаются в аппарат Гольджи для проведения повторных циклов транспорта. Вероятность возвращения гидролазы в аппарат Гольджи вместе с рецептором сильно снижается за счет удаления фосфата от маннозного остатка. Хотя существует два структурно различных маннозо-6-фосфат-ре-цепторных гликопротеина, сильно отличающихся по размерам, они имеют сходную аминокислотную последовательность и, вероятно, выполняют сходные функции.
рид при рН7 и отщепляет его при рН6; именно такой рН существует внутри эндолизосом. Лизосомные ферменты в эндолизосоме отделяются от белка-рецептора маннозо-6-фосфата и начинают расщеплять поглощенный материал, содержавшийся в эндосомах. Отделившись от «своих»
ферментов, рецепторы восстанавливают структуру и возвращаются в мембрану транс-сети Гольджи, возможно, в составе окаймленных пузырьков (рис. 8-72). Такой механизм возвращения мембран из эндолизосом обратно в аппарат Гольджи весьма напоминает круговорот мембран между эндосомами и плазматической мембраной при опосредованном рецепторами эндоцитозе (см. разд. 6.5.10). Опосредованный рецепторами транспорт лизосомных гидролаз из аппарата Гольджи к эндолизосомам аналогичен эндоцитозу внеклеточных молекул, направляющему их от плазматической мембраны в эндосомы. В обоих случаях рецепторы собираются в покрытых клатрином участках мембраны (называемых окаймленными ямками);
эти участки отшнуровываются от мембраны, образуя покрытые клатрином окаймленные пузырьки. Пузырьки доставляют затем лиганд к следующему компартменту, имеющему кислую среду, и оттуда рецепторы возвращаются в исходную мембрану.
Круговорот маннозофосфатного рецептора был прослежен с помощью специфических антител, позволяющих локализовать этот белок в клетке. В норме рецепторы маннозо-6-фосфата обнаруживают в мембранах аппарата Гольджи и эндолизосом, но не в зрелых лизосомах. Если некоторые культивируемые клетки обработать слабым основанием (например аммиаком или хлорохином), которое накапливается внутри органелл с кислой средой и поднимает там рН до нейтрального, то рецепторы исчезают из аппарата Гольджи и появляются в эндолизосомах. Можно вызвать в таких клетках возвращение рецепторов в аппарат Гольджи, либо удалив слабое основание, либо добавив в культуральную среду большое количество маннозо-6-фосфата. При обоих воздействиях рецептор отделяется от связанного с ним фермента в эндолизосоме, в одном случае в результате вторичного закисления среды в органелле, а в другом - за счет конкурентного связывания с рецептором поглощенного маннозо-6-
фосфата. Эти эксперименты свидетельствуют о том, что перемещению рецептора обратно в аппарат Гольджи способствует его конформационное изменение, связанное с отщеплением гидролазы.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
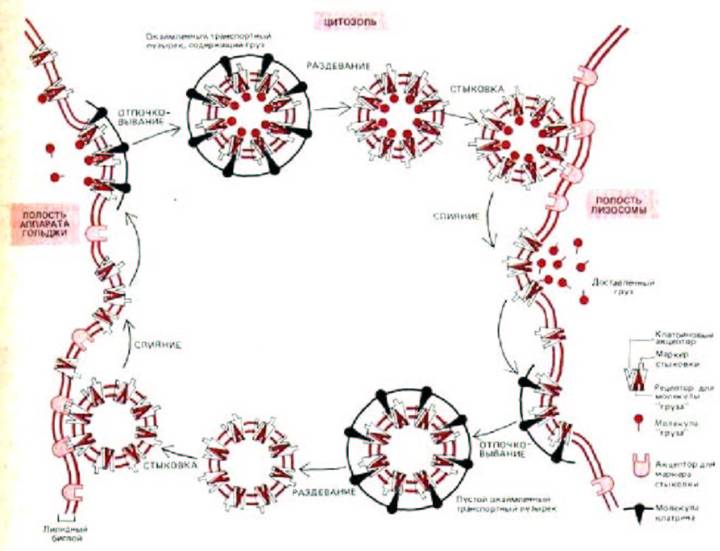
Рис. 8-73, Один из возможных механизмов, позволяющих направлять окаймленные транспортные пузырьки к определенной внутриклеточной мембране. В этой гипотетической модели молекулы «груза» - это лизосомные гидролазы, а «приемщик груза»-это белок-рецептор маннозо-6-фосфата. Изображенные здесь молекулы «маркера стыковки» и «приемщика» пока еще не охарактеризованы. Однако недавно открытые клатрин-связывающие белки, которые также связывают цитоплазматические «хвосты» некоторых мембранных белков, удовлетворяют данному здесь описанию молекул-«приемщиков». (См. Pearse, B.M., ЕМВО J. 7: 3331-3336, 1988.)
Челночная система для рецептора маннозо-6-фосфата, изображенная на рис. 8-72, является специфичной - пузырьки, несущие этот рецептор, сливаются только с нужными органеллами-мишенями, а не, к примеру, с мембранной ЭР. Считают, что клатриновая кайма на формирующихся пузырьках действует подобно «молекулярному фильтру», изолирующему рецептор и его лиганд внутри пузырьков. Однако клатрин не может отвечать за специфичность доставки пузырьков, так как кайма удаляется вскоре после их образования. Эксперименты in vitro показывают, что удаление клатрина катализируется htр70-подобным белком, а соответствующая реакция требует гидролиза АТР. Один или несколько белков, остающихся экспонированными на внешней стороне мембраны пузырька, вероятно служат специфическими «сигналами погрузки», которые узнаются комплементарным «приемщиком» на мембране органеллы-мишени.
Изучение механизмов сортировки лизосомных гидролаз весьма расширило наши представления о многих протекающих в клетках эукариот процессах переноса веществ с помощью транспортных пузырьков. Неважно, что олигосахаридные маркеры, по-видимому, больше нигде не используются, все остальные события-распознавание «груза» мембранным рецептором при отпочковании пузырька, слияние пузырька со специфичной мембраной-мишенью, высвобождение «груза» в компарт-
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
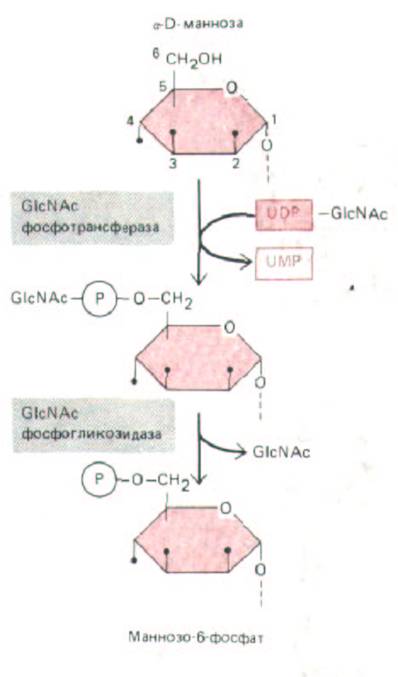
Рис. 8-74. Образование маннозо-6-фосфатного маркера на лизосомной гидролазе происходит в два этапа. Сначала GlcNAc-
фосфотрансфераза присоединяет P-GlcNA.c к 6-му углеродному атому некоторых остатков маннозьг N-связапного олигосахарида, связанного с лизосомным гликопротеином-предшественником. Затем фосфогликозидаза отрезает остаток GlcNAc и в результате образуется маннозо-6-
фосфатный маркер. Первый фермент специфически активируется сигнальным участком на лизосомных гидролазах (см. рис. 8-75), тогда как фосфогликозидаза является неспецифическим ферментом. Такая модификация некоторых остатков маннозы в цuc -компартменте Гольджи защищает их от действия маннозидаз, которые встретятся позже, в промежуточном ком-партменте аппарата Гольджи.
мент-мишень и возвращение освободившегося рецептора в исходный компартмент-вероятно, являются общими для всех видов везикулярного транспорта.
[58]
Система отделения лизосомных гидролаз и отправки их в эндолизосомы работает благодаря тому, что маннозофосфатные группы добавляются в аппарате Гольджи только к определенным гликопротеинам. Для этого требуется специфическое узнавание гидролаз ферментами аппарата Гольджи, ответственными за присоединение маннозо-6-фосфата (М6Ф). Поскольку все гликопротеины, попадающие в компартмент Гольджи, имеют идентичные N-связанные олигосахаридные цепи, сигнал, по которому к олигосахариду добавляется М6Ф, должен находиться в самой полипептидной цепи каждой гидролазы.
Для присоединения маннозофосфатных групп к лизосомным гидролазам необходима последовательная работа двух ферментов. N-
ацетилглюкозамин-фосфотрансфераза (GlcNAc-фосфотрансфераза) переносит GlcNAc-P-часть молекулы нуклеотид-сахара UDP-GlcNAc к остатку маннозы в олигосахарпде, а второй фермент - фосфогликозидаза-затем удаляет концевой GlcNAc, оставляя фосфат, в результате образуется маннозо-6-фосфат М6Ф (рис. 8-74). Фосфотрансфераза специфически связывается с гидролазой благодаря участку (или сайту) узнавания, не совпадающему с активным центром этой реакции (рис. 8-75). По-видимому, в данном случае сигнал для сайта узнавания представляет собой сигнальный участок, а не сигнальный пептид (см. разд. 8.1.6). Об этом свидетельствует тот факт, что при частичном
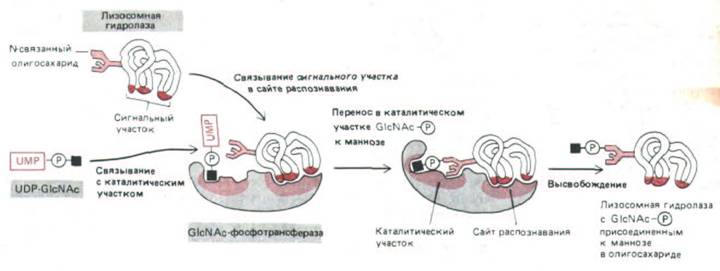
Рис. 8-75. У фермента GlcNAc-фосфотрансферазы, узнающей лизосомные гидролазы в аппарате Гольджи, участок узнавания и активный центр не совпадают. В активном центре связываются богатые маннозой N-связанные олигосахариды и UDP-GlcNAc. Сайт узнавания объединяется с сигнальным участком, имеющимся только на поверхности лизосомных гидролаз и их предшественников.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
разворачивании гидролазы распознавание практически прекращается.
Как только фосфотрансфераза узнает сигнал на гидролазе, она присоединяет GlcNAc-P к одному или двум маннозным остаткам в каждой олигосахаридной цепи. Поскольку большинство лизосомных гидро-лаз несут многочисленные олигосахариды, они приобретают много остатков М6Ф, что приводит к многократному усилению сигнала. Если при связывании обычной лизосомной гидролазы с сайтом узнавания фосфотрансферазы Ка составляет 105 л моль, то для множественно фосфорилированной гидролазы и маннозофосфатного рецептора соответствующий показатель равен 104 л/моль, т.е. их связь в 10000 раз прочнее.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2160; Нарушение авторских прав?; Мы поможем в написании вашей работы!