КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Проникнення йонів крізь мембрану. Мембранний потенціал спокою
|
|
|
|
Проникністю біологічних мембран називають їхню здатність пропускати у клітину або випускати з неї йони та молекули різних речовин.
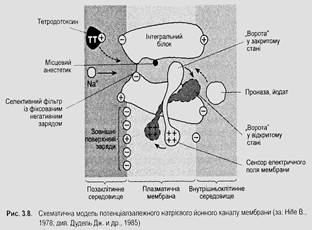
Проникнення йонів крізь мембрану за градієнтами концентрації названо пасивним транспортуванням. Для полегшеної дифузії йонів крізь мембрану слугують йонні канали. Вони є інтегральними білками. Йонні канали неоднорідні. Для проникнення кожного виду йонів (ІС, СІ і Са") є окремий тип каналів. Навіть для проникнення одного і того ж виду йонів у спокої та при збудженні слугують різні канали. Розрізняють канали витоку (для проникнення йонів у спокої), потенціалзалежні та хемочутливі йонні канали.
t Канали витоку забезпечують проникнення йонів у стані спокою.
t Потенціалзалежні йонні канали у спокої закриті й не пропускають йонів. Вони відкриваються (активуються) у відповідь на деполяризацію мембрани і забезпечують проникнення йонів під час генерування потенціалів дії.
t Хемочутливі йонні канали також закриті у стані спокою, активуються за дії медіаторів, гормонів, внутрішньоклітинних посередників та інших фізіологічно активних речовин і забезпечують проникнення йонів під час генерування постсинаптичних потенціалів, фото- і хеморецепції тощо. Отже, у стані спокою відкриті тільки канали витоку, які позбавлені ворітних механізмів. Вони забезпечують проникнення йонів під час генерування мембранного потенціалу спокою.
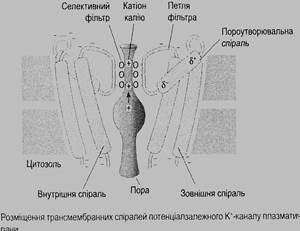
Для вибіркового проникнення йонів канали витоку мають селективні фільтри. Потенціалзалежні (рис. 3.8) та хемочутливі канали мають і активаційні ворота, які у стані спокою закриті.
Стверджують (В. Ґанонґ 2002), що у ссавців є понад 40 різноманітних калієвих каналів і понад 30 натрієвих та кальцієвих.
Оскільки через йонні канали проникають йони, їхній рух зумовлює появу струму. Здатність каналів проводити струм характеризують провідністю. Цей струм можна виміряти методом фіксації потенціалу в умовах внутрішньоклітинної перфузії та використати для розрахунку провідності (О, вимірюють у сименсах, См). Для секреторних клітин у стані спокою характерне таке співвідношення провідностей:
Gk+: GNa+: Gcl- = 1:0,08:0,28
Мембрана характеризується у спокої різною проникністю для йонів завдяки різній провідності каналів: найбільш проникна вона для К+, менше — для СІ-, ще менше - для Na+; Са2+ у стані спокою практично не проникає у клітини.
Проникність мембрани для кожного з йонів оцінюють за коефіцієнтами проникності (Р, вимірюють См/с), які для мембрани гігантського аксона кальмара співвідносяться між собою як:
Pk+ : PNa+: Pcl- = 1:0,04: 0,45
Оскільки мембрана у спокої найбільш проникна для йонів К+, деяка кількість їх дифундує з клітини і зосереджується ззовні плазматичної мембрани, де утримується електростатично внутрішньоклітинними органічними аніонами, які не можуть покинути клітину разом з К+. Так створюється розділення позитивних і негативних зарядів по обидва боки мембрани, тобто виникає мембранна різниця потенціалів.
Ë Між зовнішньою і внутрішньою поверхнями плазматичної мембрани клітин наявна різниця потенціалів, яка у стані спокою (тобто у незбуджених і незагальмованих клітинах) становить переважно -60...-90 мВ і називається мембранним потенціалом спокою.
Мембранний потенціал спокою фактично характеризує величина потенціалу внутрішньої поверхні мембрани, яка відносно землі (нульовий потенціал) заряджена негативно. У скелетних м'язових волокнах мембранний потенціал спокою досягає -95 мВ, у кардіоміоцитах не перевищує -85 мВ, у клітинах гладеньких м'ячів досягає -60 мВ, у нервових клітинах і волокнах не перевищує -80 мВ. у секреторних клітинах лежить у межах від -20 до -60 мВ.
Наявність мембранного потенціалу характерна особливість усіх живих клітин, з їхнім відмиранням він зникає. Причинами його виникнення є нерівномірний розподіл йонів між цитоплазмою та міжклітинним середовищем і вибіркова проникність мембрани для різних іонів.
Різницю потенціалів, яка виникає на мембрані за рахунок дифузії К+ назовні, можна розрахувати за формулою Нернста:
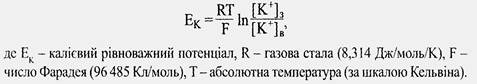
Мембранна різниця потенціалів, яка генерується К+, може модифікуватися за рахунок дифузії крізь мембрану Na+ та С1-. Проникнення Na+ у клітини спричинює деяке зменшення мембранного потенціалу, або деполяризацію мембрани. Дифузія СІ- у клітини, навпаки, забезпечує деяке збільшення трансмембранної різниці потенціалів (гіперполяризація).
Підсумкову величину мембранного потенціалу, що генерується за рахунок дифузії К+, Na+, СІ-, можна розрахувати за формулою Ґольдмана:

Отже, основна частина мембранного потенціалу спокою, яку називають концентраційним потенціалом, формується за рахунок дифузії йонів, переважно К+ крізь мембрану.
Відомо, що на кожний йон К+, який транспортується натрій-калієвою помпою в клітину, з неї виводяться три йони Nа+. За рахунок цього у клітині створюється дефіцит позитивних або надлишок негативних зарядів. Тобто деяка різниця потенціалів генерується прямо помпою і додається до концентраційного потенціалу. У цьому разі помпа характеризується елєктрогенним ефектом. Тому натрій-калієва помпа у формуванні величини мембранного потенціалу спокою виконує дві функції: підтримує трансмембранні градієнти Na+ і К+ та прямо генерує деяку різницю потенціалів.
Отже, реальний мембранний потенціал складається з концентраційного потенціалу та електрогенного ефекту натрій-калієвої помпи. Внесок електрогенного ефекту помпи у мембранний потенціал не перевищує -15 мВ. Його можна швидко ліквідувати строфантином, охолодженням або чинниками, що ведуть до дефіциту АТФ.
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 2299; Нарушение авторских прав?; Мы поможем в написании вашей работы!