КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Структура и функции митохондрий. Генетическая система митохондрий. Онтогенетический и филогенетический аспекты митохондриогенеза.
|
|
|
|
В растениях митохондрии впервые были идентифицированы световой микроскопией как частицы, которые связывали краситель "Янус зеленый Б". Изолированные митохондрии растений обычно сферической или продолговатой формы диаметром 0,5—1,0 мкм и длиной до 3 мкм. Много митохондрий содержится в замыкающих клетках устьиц. Количество митохондрий у растительных клеток меньше, чем в типичной животной клетке. Ультраструктура митохондрий растений похожа на строение митохондрий других объектов.
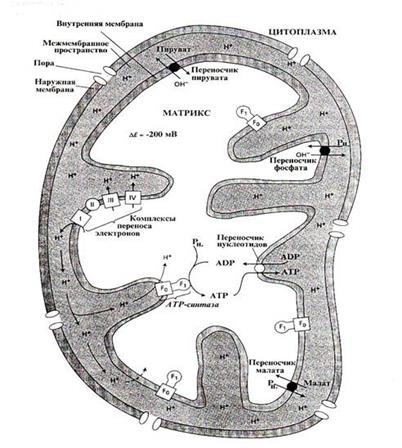
Митохондрии имеют две мембраны: гладкую наружную и внутреннюю, образующую многочисленные выросты — кристы. Пространство между наружной и внутренней мембранами называется межмембранным (перимитохондриальным). Оно является внешней средой для внутренней мембраны и матрикса митохондрий. В растительных митохондриях осуществляются процессы аэробного дыхания, окислительного фосфорилирования, превращения ряда аминокислот, синтез жирных кислот и некоторых фосфолипидов. Интактные митохондрии осмотически активны: способны набирать и отдавать воду соответственно в гипо- и гиперосмотической среде. Большинство неорганических ионов и заряженных органических молекул не способны диффундировать в матрикс. Осмотическим барьером является внутренняя мембрана митохондрий. Наружная мембрана хорошо проницаема для соединений, молекулярная масса которых ниже 10 кДа, т. е. для большинства ионов и метаболитов. Подобно хлоропластам митохондрии являются полуавтономными органеллами, поскольку содержат рибосомы, РНК и ДНК, а также ферменты и кофакторы, необходимые для синтеза белков, кодируемых митохондриальной ДНК. Способность ДНК митохондрий к репликации позволяет этим органеллам делиться независимо от деления ядра, поэтому они пролиферируют делением, а не образованием denovo. Липидная часть обеих мембран состоит главным образом из фосфолипидов, 80% которых — фосфатидилхолин и фосфатидилэтаноламин.
Многие особенности молекулярной генетики растительных митохондрий сходны с другими эукариотическими организмами. ДНК митохондрий является кольцевой молекулой и наследуется по материнской линии. Митохондриальный геном растений кодирует большинство тРНК, необходимых для синтеза митохондриальных белков, ряд рибосомальных белков и некоторые элементы электрон-транспортной цепи. Однако большая часть митохондриальных белков, включая ферменты цикла Кребса, кодируется ядерными генами и импортируется в митохондрии из цитоплазмы. В растениях, в отличие от животных, простейших и грибов, митохондриальный геном имеет ряд отличительных особенностей. Хотя митохондриальный геном высших растений кодирует только около 50 различных белков, он крупнее, чем у животных клеток, и сильно варьирует по величине у различных видов растений (от 218 т.п.н. у дикой редьки до 570 т.п.н. у кукурузы). Еще одной удивительной особенностью растений является тот факт, что нуклеотидная последовательность митохондриальной ДНК не комплементарна мРНК, образующейся при транскрипции. Тем не менее потеря комплементарности при передаче генетической информации не сказывается на качестве генных продуктов — белков, кодируемых митохондриальной ДНК. Это обеспечивается исправлением (editing) "некомплементарных" мРНК перед трансляцией. Процессы исправления "некомплементарных" мРНК обнаружены в митохондриях многих организмов, однако у растений это явление встречается чаще. Большинство исправлений сводится к окислению аминогруппы у 6-углеродного атома цитозина и превращению его в урацил.
** клетка съела клетку àмитохондрия
*может делиться вне зависимости от деления самой клетки
И др. факты из жизни митохондрий см. прошлые года
28. Пути окисления глюкозы при дыхании. Гликолиз, пентозофосфатный окислительный путь. Последовательность реакций, ферменты, энергетический выход.
Главным дыхательным субстратом являются углеводы. В растительной клетке существует даже такая закономерность: чем больше она содержит сахаров, тем интенсивнее она дышит. Окисление дыхательного субстрата - сложный процесс, состоящий из большого числа химических реакций. Главный путь распада углеводов - гликолиз и ЦТК (ц. Кребса).
Гликолиз(дихотомический распад углеводов)
Гликолиз – это последовательность реакций, приводящих к превращению гексозы (глюкозы) в пируват (ПВК) с одновременным образованием АТР. Гликолиз – процесс анаэробный, но он может протекать и в отсутствии кислорода, и в присутствии О2. Гликолиз является основным метаболитическим путем, образующим энергию в форме АТР в клетках, где отсутствует фотосинтез. Все реакции гликолиза протекают в цитоплазме.
Гликолизу предшествует гидролиз или фосфоролиз сложных углеводов до глюкозы, которая и вступает в первую реакцию гликолиза, а после гликолиза протекает аЭробное дыхание или аНАэробное брожение. Если гидролиз – фермент 3 класса, если фосфоролиз – ф.2 класса, в этом случае должна протекать еще одна реакция – превращение глюкозо-1-фосфата в глюкозо-6-фосфат под действием фермента 5 класса фосфоглюкомутазы.
Процесс гликолиза можно разделить на два этапа. Первый этап, протекающий с расходом энергии 2-х молекул АТФ, заключается в расщеплении молекулы глюкозы на 2 молекулы глицеральдегид-3-фосфата.
На втором этапе происходит окисление глицеральдегид-3-фосфата, сопровождающееся синтезом АТФ.
Первый этап гликолиза (эндергонический) начинается с реакции фосфорилирования глюкозы и требует затраты химической энергии (в форме АТР), фосфорилированная форма глюкозы является наиболее реакционно активной. Первый этап включает в себя 5 реакций, две из которых (1-ая и 3-ья являются НЕобратимыми).
Описание реакций 1го этапа:
1 реакция: Глюкоза превращается в глюкозо-6-фосфат, при этом АТР переходит в АДР, фермент – 2 класса, гексокиназа (глюкокиназа). Реакция необратимая. Р. Фосфорилирования.
2 р. Глюкозо-6-фосфат превращается в фруктозо-6-фосфат, фермент – 5 кл. Фосфоглюкоизомераза. Р. Обратима. Р. Изомеризации.
3 р. Второе фосфорилирование. Фруктозо-6-фосфат под действием АТР переходит во фруктозо-1,6-бисфосфат. Фермент – 2 кл. Фосфофруктокиназа. (играет важную роль в регуляции скорости гликолиза). Р. НЕобратима.
4 р. Расщепления фруктозо-1,6-бисфосфата под действием фермента 4 класса альдолазы, превращается в ФГА и ДОАФ. Р.Обратима.
5 р. Взаимопревращение ФГА и ДОАФ, фермент 5 кл. триозофосфатизомераза.
Во втором этапе участвует ФГА.
|
|
|
|
|
Дата добавления: 2017-02-01; Просмотров: 103; Нарушение авторских прав?; Мы поможем в написании вашей работы!