КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Исследование нуклеиновых кислот. Методы ДНК-диагностики 3 страница
|
|
|
|
· амплифицируйте у каждого члена семьи полиморфный аллель с помощью праймеров Р1 и Р2,
· пометьте амплифицированные продукты с помощью радиоактивной или флюоресцентной метки,
· гибридизуйте каждый продукт с микрочипом и выявите квадрат, в котором произошло связывание с меченой пробой. Этот квадрат будет показывать, какой аллель присутствует у данного члена семьи.
Технология может быть использована для анализа полиморфизма генома. В частности, 149 микрочипов, каждый из которых содержал 150 000— 300 000 олигонуклеотидов, были использованы для скрининга 263 млн п. о. последовательности генома человека, и в результате был обнаружен 3241 полиморфный сайт. Следует обратить внимание на то, что во всех случаях исследовалась определенная, небольшая по сравнению с полным геномом, область.
Несмотря на впечатляющие результаты и кажущуюся простоту описанной выше технологии, использующей олигонуклеотидные микрочипы, существует множество проблем, связанных с ее практическим использованием. Микрочипы могут давать высокий уровень ложнопозитивных ответов при обнаружении SNP. Инсерции, вставки, делеции и перегруппировки трудно поддаются анализу этим методом. Повторяющиеся элементы генома также способны вносить серьезные осложнения в анализ.
Помимо детекции мутаций и полиморфизмов, олигонуклеотидные микрочипы предполагают использовать для создания генетической карты третьего поколения. В частности, олигонуклеотиды, соответствующие SNP, легко можно поместить на микрочипы. Поскольку SNP в основном диаллельны, для создания карты их нужно раза в 3 больше, чем мультиаллельных маркеров. Можно предвидеть множество других применений олигонуклеотидных микрочипов, включая анализ сцеплений и ассоциаций, потерю гетерозиготности и ДНК-дактилоскопию.
Другой важный аспект использования олигонуклеотидных микрочипов — анализ экспрессии генов. Анализ особенностей экспрессии генов — один из необходимых элементов для понимания их функций. 20-звенный олигонуклеотид, представляющий определенный ген, по-видимому, достаточен для надежного обнаружения продукта этого гена среди клеточных РНК. Содержание транскриптов может быть измерено вплоть до нескольких копий на клетку. Удается прослеживать экспрессию 10 000 генов на 1 чип. Эта система уже использована для изучения изменений в экспрессии генов в раковых клетках. Сравнение экспрессии 900 генов в 2 клеточных линиях показало, что при опухолевой трансформации 1,7% генов понижало уровень экспрессии, тогда как 7% — повышало.
Для анализа экспрессии генов используют также другой вариант микрочипов, содержащих не олигонуклеотиды. а иммобилизованные кДНК. В случаев кДНК-микрочипов ППР-амплифицированные фрагменты индивидуальных кДНК наносят в разные квадраты микрочипа, так что каждый квадрат ставится в соответствие с определенным геном. ДНК фиксируют в этих квадратах и денатурируют, чтобы сделать ее одноцепочечной. Чип с иммобилизованной ДНК гибридизуют со смесью различно флюоресцентно меченных кДНК, полученных из сравниваемых тканей, точно так же, как это делалось в случае олигонуклеотидных микрочипов.
Недавно кДНК-микрочипы использовали также для анализа различий близкородственных геномов. В этом случае технология была использована для анализа ДНК из клеточных линий и опухолей с известными хромосомными аномалиями, чтобы оценить перспективность метода. Показана возможность обнаружения делеции в одной из хромосом и различия в числе копий хромосом в анеуплоидных клетках.
Хотя исходной идеей использования микрочипов было быстрое секвенирование ДНК, сейчас эта идея отошла на задний план. Однако чипы могут быть использованы для повторного секвенирования уже известных последовательностей. В частности, был ресеквенирован полный митохондриапьный геном длиной 16,6 килобаз, митохондриальный чип содержал 13600025-меров. Полная последовательность была прочитана с 99% точностью, что позволяет осуществить однозначное определение митохондриальных мутаций. Скорость такова, что полная митохондриальная геномная последовательность может быть получена за 12 мин. В настоящее время этот чип используется для изучения вариабельности митохондриальной ДНК в популяциях. Исследуется возможная роль митохондриальных мутаций в нервологических заболеваниях. Понятно, что возможны и другие применения ресеквенирования для анализа известных участков генома.
В частности, микрочипы в усовершенствованном варианте будут использоваться в будущем для анализа природы сложных болезней, который требует полногеномного подхода.
Микрочипы принципиально открывают возможность сравнительного анализа экспрессии практически всех генов в различных клетках. Появляется путь к исследованию влияния громадного разнообразия генетических факторов и факторов окружающей среды на изменения экспрессии. Создаются предпосылки для анализа тотальных изменений экспрессии генома в разных типах клеток при болезнях. Это в свою очередь создает новые возможности для функционального анализа и диагностики.
В изделиях, которые промышленность выпускает для нашего повседневного обихода, используется около 80 тысяч различных химических веществ. И, как ни странно, подавляющее большинство из них никогда полностью не проверялось на вредность. Издавна всякое новое вещество проверяют на животных. Но дело это хлопотное, долгое и дорогое, да и защитники животных, случается, суют палки в колеса. Но самое главное: данные, полученные на животных, не всегда годятся для людей. Так что, стопроцентной уверенности в безвредности того или иного лекарства, пищевой добавки, косметического средства и других веществ, при всем желании, не получается.
Выход из положения сулит применение метода, который предлагает новое научное направление - токсикогеномика. Животные клетки в специальной пробирке входят в соприкосновение с проверяемым веществом. Собственно, это даже не клетки, а так называемые ДНК-чипы. Входящие в ДНК гены реагируют на вещество: одни гены активизируются, другие, наоборот, выключаются, и по этой генной реакции нетрудно определить, безвредно вещество, или нет.
Преимущества нового метода очевидны - говорят его создатели. Традиционные испытания веществ на животных, могут показать, что именно в них действует разрушительно на печень, что вызывает рак или приводит к врожденным физическим недостаткам. Но почему это происходит - останется загадкой. Разрешит ее лишь анализ генной активности.
Новый метод может значительно уменьшить количество подопытных животных - ведь он, в основном, ориентируется на клетки. Правда, в отдельных случаях генная активность проверяется и на живых животных, бывает также необходимо дополнить новый метод традиционным. Но, в целом, тенденция здесь клеточная. Еще один плюс в том, что генная активность может сигнализировать об опасности задолго до появления ее видимых симптомов, например, до появления опухоли. Генетический уровень чувствительнее физиологического. О скорости метода в сравнении с традиционной проверкой на животных, по мнению его сторонников, и говорить не приходится: это преимущество не нуждается в доказательствах.
Однако фармацевтические и химические фирмы относятся к новинке со смешанным чувством. Их представители говорят, что результаты испытаний, проведенных по генетической методике, могут, как это ни парадоксально, быть истолкованы неверно или пристрастно, чем тут же воспользуются защитники окружающей среды и начнут добиваться введения запрета на вполне невинный продукт. «Внедрение в практику ДНК-чипов откроет собой эпоху бесконечных судебных процессов, на которых создатели новых веществ будут терпеть поражение за поражением», - предсказывают специалисты.
«Придраться можно ко всему, особенно, в нашей области, - говорит Джозеф Сайна, токсиколог одной из фармацевтических фирм и редактор журнала «Молекулярная токсикология». - Только бы новый метод не поторопились использовать до того, как он будет доведен до совершенства - тогда ошибок не избежать. Ложные тревоги поведут к неоправданным запретам, а запоздалые тревоги и вовсе скомпрометируют новшество».
Выяснить, какие гены активизируются от контакта с новым веществом, с помощью ДНК-чипов несложно. Когда ген, состоящий из ДНК, становится активным, он создает молекулу РНК, которая сообщает клетке, какой именно белок ей надлежит выработать. Фиксируя эту информационную РНК, ученые видят, какие гены активизировались.
ДНК-чип представляет собой микроскопическую пластинку из стекла или пластика, содержащую тысячи генов. Как различить, что на ней происходит? Обычно информационная РНК из клетки снова превращается в ДНК, после чего окрашивается светящейся краской. Каждый кусочек этой ДНК прилепляется к соответствующей ДНК на чипе, эта точка светится, как светятся и сотни других. Так на чипе возникает паттерн, или рисунок генной активности.
Подобные чипы уже широко применяются. Например, в Боулдере разрабатывают специализированные чипы для онкологии, где сравнивают генную активность в раковых и здоровых клетках. О генной активности будет по-прежнему свидетельствовать мозаика светящихся точек.
Но можно ли истолковать эту мозаику однозначно? Ведь гены в процессе работы организма могут включаться и выключаться по самым разнообразным причинам. В одном случае на генной активности отразится естественная гибель клеток, в другом - это будет реакция на нейтральное вещество. И так далее. «Множество перемен в активности генов не связано с токсичностью, - указывает Крис Кортон из Института токсикологии в Северной Каролине. - Вспомним хотя бы ген, ведающий ферментом, разрушающим многие лекарства и химические вещества. Этот ген включается от углекислого газа, от брюссельской капусты и еще, Бог знает от чего. Фармацевтические фирмы всегда опасаются его неожиданных реакций».
Невзирая на эти сложности, ученые испытывают новым методом сотни веществ с заведомой токсичностью, чтобы пополнить базу генетических данных, с которыми можно будет сравнивать картину, получаемую от веществ с неизвестными свойствами. В США начал работать недавно организованный Центр токсикогеномики, в задачу которого входят как фундаментальные исследования, так и создание такой базы данных, которой могли бы безвозмездно пользоваться все, кто пожелает.
Сознавая многие несовершенства нового метода, ученые говорят, что, в принципе, он работает. Им удалось найти вполне определенные паттерны для таких токсических соединений, как разрушители гормонов, канцерогены и опасные для печени яды. Фирма «Джонсон энд Джонсон» установила, как вели себя гены, когда клетки печени входили в контакт с сотней различных токсичных веществ. Не зная, что это за вещества, компьютер сумел сгруппировать их по типу токсичности. Результаты этой работы опубликованы в последнем выпуске журнала «Токсикологические науки». Другая фирма - «Берринген Ингельхайм» - использовала ДНК-чипы для определения таких типов токсичности, которые в обычных условиях потребовали бы длительных исследований клеток с помощью электронного микроскопа.
Интересное и полезное исследование провела фирма «Phase 1», с которой, кстати, сотрудничают многие из упомянутых фармацевтических компаний. Она обнаружила 260 генов, проявляющих специфическую активность у людей с аллергией на пенициллин. Нередко бывает так, что при клинических испытаниях нового лекарства, прошедшего перед этим проверку на животных, пациенты страдают от побочных эффектов. Теперь, как говорит Спенсер Фар - глава фирмы «Phase 1», можно будет предвидеть генетический ответ пациентов на новое лекарство и не тратить миллионы попусту. Фар убежден, что у нового метода блестящие перспективы.
Разрабатываются и другие методы, позволяющие кардинально улучшить проверку веществ на животных. Один из них, в частности, предлагает генная инженерия. Кроме того, токсикогеномика пока еще дороговата. Один ДНК-чип стоит 1000 долларов. Правда, чипы будут постоянно дешеветь.
Чипы могут и не заметить опасность. Например, побочный эффект от нового лекарства может возникнуть из-за его взаимодействия с другим лекарством. Сложные взаимодействия веществ чипы пока не улавливают. Поводов для сомнений в новом методе почти столько же, сколько и поводов для уверенности в его преимуществах. Так что, обсуждение перспектив токсикогеномики пока в самом разгаре.
Молекулярно – генетические методы исследований в судебной медицине. Прогресс молекулярно-биологической науки открыл новые пути решения проблемы судебно-медицинской идентификации личности, обеспечив возможность выявления индивидуализирующих личность признаков на уровне не фенотипа, а генетической матрицы - клеточной ДНК. Молекулярно-генетические маркерные системы основаны на существовании различий в структуре ДНК (генов) у разных индивидуумов. Гомологичные гены, то есть те, что определяют формирование одного и того же признака, например, форму носа или цвет глаз, у разных людей могут находиться в разных аллельных состояниях. На молекулярном уровне аллельные варианты одного и того же гена отличаются небольшими изменениями в структуре их ДНК, конкретно, в последовательности нуклеотидов в полинуклеотидной цепи. Возможны замена единичных нуклеотидов, так называемые точковые замены, или локальные перестройки, именуемые делециями и инсерциями - соответственно утрата либо добавление небольших участков цепи. Столь незначительные различия в конечном итоге и определяют то, чем разные люди отличаются друг от друга: уникальное сочетание аллельных вариантов всех генов обеспечивает биологическую индивидуальность каждого человека.
Геном человека содержит десятки тысяч генов и состоит из более чем 3 млрд. нуклеотидных пар, при этом молекулы ДНК любых двух людей (неродственников) отличаются в среднем только одним нуклеотидом из каждых трехсот-четырехсот. Но даже такие отличия, как правило, носят характер случайных отклонений от некой доминирующей нормы. Теоретически это означает, что если у сотни человек проанализировать фрагмент ДНК длиной 300-400 нуклеотидов для одного и того же среднестатистического гена, то девяносто девять человек вполне могут оказаться неотличимыми друг от друга. Практическое значение для целей генетической индивидуализации личности (как предпосылки для ее идентификации) имеют отнюдь не любые гены, а только такие, у которых много аллельных форм.
На практике в качестве маркеров индивидуальности выступают мультиаллельные гипервариабельные гены (гипервариабельные генетические локусы). Индивидуализирующими характеристиками служат многочисленные структурные варианты таких локусов, которые в разных сочетаниях присутствуют в ДНК разных индивидуумов. Открытие в начале 1980-х годов феномена локального генетического гиперполиморфизма в лаборатории Р. Уайта в США и позже разработка А. Джеффрисом высокоэффективных молекулярных зондов типа минисателлитной ДНК предоставили новые, недостижимые ранее возможности для решения проблемы индивидуализации человека и установления кровнородственных связей.
Технологии молекулярно – генетической индивидуализации. В судебно-экспертной практике базовыми молекулярно-генетическими технологиями признаны: анализ полиморфизма (вариабельности) длины рестриктазных фрагментов ДНК, анализ полиморфизма длины амплифицированных фрагментов ДНК и анализ полиморфизма нуклеотидных последовательностей (сайт-полиморфизма) ДНК.
Анализ полиморфизма длины рестриктазных фрагментов ДНК. Большая часть вариаций в полинуклеотидной цепи геномной ДНК вызвана точковыми нуклеотидными заменами и некоторыми другими вариантами реорганизации нуклеотидных последовательностей - инверсиями, делециями и инсерциями. В результате в молекулах ДНК появляются новые или утрачиваются существовавшие ранее участки воздействия (сайты) особых ферментов - рестрикционных эндонуклеаз (рестриктаз), в которых они расщепляют полинуклеотидные цепи ДНК.
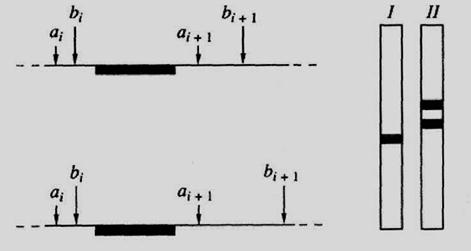
Рис.52. Схема блот-гибридизациии фрагментов ДНК
Слева - схема расположения на гомологичных хромосомах соседних участков узнавания (аі и ai+i; bi и bi+i) двух рестриктаз (I и II), зачерненный участок - область гомологии зонда; справа - схематическая картина результата блот-гибридизации с данным зондом суммарной ДНК, гидролизованной рестриктазами I и II, обусловливает изменение длины получающихся рестриктазных фрагментов ДНК.
Интересующие фрагменты ДНК можно визуализировать путем молекулярной гибридизации с соответствующим зондом; данный метод хорошо известен в молекулярной биологии нуклеиновых кислот как блот-гибридизационный анализ (рис. 52). С его помощью полиморфные участки генома обнаруживаются в виде имеющих разную длину гомологичных фрагментов ДНК, которые образуются после гидролиза геномной ДНК рестриктазами. Этот феномен получил название полиморфизма длины рестриктазных фрагментов (ПДРФ) ДНК.
Индивидуальный полиморфизм длины рестриктазных фрагментов создает предпосылки для решения задач, связанных с индивидуализацией организма и установлением его биологических родственных связей с другими индивидуумами. Однако большинство полиморфных геномных локусов - потенциальных генетических маркеров - имеет только два варианта: дикий тип/мутация, то есть являются диморфными или диаллельными. Ценность диморфных маркеров невелика, потому что у многих людей может оказаться один и тот же вариант такого гена. Поэтому, как уже указывалось, индивидуализирующее значение имеют не любые, а только гипервариабельные полиморфные фрагменты.
Поскольку гипервариабельные локусы мультиаллельны, то информативность маркерных систем на их основе намного выше, чем информативность систем (как правило, диаллельных), базирующихся на единичных нуклеотидных заменах. Теоретически можно предполагать, что высокая локальная генетическая вариабельность вызвана не столько точковыми нуклеотидными мутациями или микроделециями/инсерциями, сколько другими механизмами, приводящими к более существенной реорганизации геномных последовательностей: транспозициями, неравными или незаконными рекомбинациями, проскальзыванием репликативного комплекса и т.п. Как оказалось, такие локусы достаточно распространены в геноме человека.
Наиболее примечательны из них «минисателлитные» ДНК, впервые описанные Джеффрисом. Это относительно короткие (10-60 нуклеотидных пар), рассеянные по геному, повторяющиеся нуклеотидные последовательности, имеющие тандемную организацию и демонстрирующие разную степень внутригрупповой гомологии. Число тандемных повторов в минисателлитных блоках (и следовательно, длина самих блоков) варьирует в широких пределах - от трех-четырех до нескольких тысяч; такие блоки представляют разные аллельные варианты мультиаллельных генетических локусов.
В 1987 г. И. Накамура предложил называть такого рода генетические элементы локусами с варьирующимся числом тандемных повторов (рис.53), или просто тандемными повторами с переменным числом звеньев (общепринятая английская аббревиатура VNTR - Variable Number Tandem Repeat).
Вариации числа повторяющихся элементов в VNTR-блоках как раз и обусловливают структурный полиморфизм этих локусов, проявляющийся в форме ПДРФ. Дело в том, что длина соответствующих рестриктазных фрагментов, если отсутствуют участки расщепления внутри самих повторов, зависит от числа повторяющихся единиц.
Пожалуй, наиболее значимое свойство открытых Джеффрисом минисателлитов - их существование в виде семейств, диспергированных по геному. Все локусы каждого семейства содержат наборы тандемных олигонуклеотидных последовательностей, объединяемых одним нуклеотидным «мотивом», или кор-последовательностью. Таким образом, помимо мультиаллельности, то есть полиморфизма, обусловленного переменным числом повторяющихся элементов в самих локусах, минисателлитные маркеры
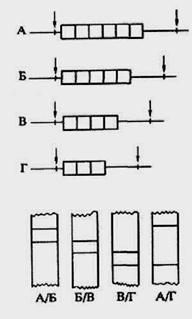
Рис.53. Последовательности ДНК с варьирующимся числом тандемных повторов: вверху - варианты таких локусов А, Б, В и Г различаются по ширине на один повторяющийся сегмент, сайты рестрикции - мечены стрелками; внизу - схематическая картина блот- гибридизации с молекулярным зондом, гомологичным идейному повтору, некоторых гетерозиготных по данному локусу ДНК.
характеризуются еще и «системным» полиморфизмом - возможностью одновременного выявления аллельных вариантов сразу нескольких (часто генетически не сцепленных между собой) локусов, представляющих одно семейство. В результате информативность таких гиперполиморфных мультилокусных систем оказывается предельно высокой, в принципе, достаточной для доказательной индивидуализации человека. В настоящее время неизвестно общее число семейств гипервариабельных локусов в геноме человека, но, по-видимому, оно достаточно велико; каждое из них потенциально представляет собой мультилокусную гипервариабельную систему с очень высоким индивидуализирующим потенциалом.
Анализ полиморфизма длины амплифицированных фрагментов ДНК. Не требующий использования рестрикционных ферментов и отличающийся чрезвычайно высокой чувствительностью, подобный анализ стал возможным в результате внедрения в практику метода так называемой энзиматической амплификации гипервариабельных генетических локусов с помощью полимеразной цепной реакции (ПЦР). Сейчас области применения этой реакции, как в фундаментальной науке, так и в биотехнологии столь разнообразны и многочисленны, что трудно их даже перечислить. Главный же смысл этого открытия в том, что удалось преодолеть все ограничения молекулярно-генетических методов исследования, которые были связаны с недостаточным для анализа количеством ДНК.
Полимерная цепная реакция позволяет выделить и размножить любую необходимую для анализа последовательность ДНК в количестве, превышающем исходное в десятки и даже сотни миллионов раз. Такая высокая степень направленного обогащения теоретически позволяет работать с единичными молекулами ДНК, следовательно, открывается возможность молекулярно-генетического типирования даже в случае исчезающе малых количеств доступного для экспертизы биологического материала. Действительно, еще в 1986 г. Джеффрис экспериментально доказал, что с помощью ПЦР реальной становится задача геномной идентификации на уровне нескольких или даже одной клетки. Кроме того, ПЦР весьма эффективна при анализе сильно разрушенных ДНК, подвергшихся высокой степени деградации.
Метод энзиматической амплификации ДНК положен в основу создания высокоспецифичных диагностических и индивидуализирующих тест-систем. Все они разрабатываются по единому принципу, который заключается в подборе праймеров на известной последовательности нуклеотидов ДНК (как правило, это олигонуклеотиды длиной 20-25 нуклеотидов, которые располагаются по флангам высокополиморфных мультиаллельных локусов) и оптимизации условий энзиматической амплификации выбранных генетических локусов.
Как мы уже знаем, гипервариабельные локусы VNTR-типа содержат различное число повторяющихся элементарных звеньев. При амплификации с праймерами, расположенными по краям такого локуса, образуются фрагменты ДНК различной длины, причем длина фрагмента будет пропорциональна числу повторов. Таким образом, в данной аналитической системе любой индивидуальный образец ДНК человека характеризуется наличием двух амплифицированных фрагментов разной или одинаковой длины (соответственно, гетерозиготное и гомозиготное состояние), поскольку каждая из двух гомологичных хромосом несет свой вариант гипервариабельного локуса, который может отличаться от других числом содержащихся в нем тандемных повторов. Этот феномен получил название полиморфизма длины амплифицированных фрагментов (ПДАФ) ДНК.
Молекулярные копии амплифицируемого локуса ДНК накапливаются в реакционной смеси, вследствие чего количественно оказываются доступными для сравнительного анализа. Далее полученные амплификационные продукты фракционируют с помощью электрофореза в геле агарозы или полиакриламида, фрагменты ДНК, амплифицированные в ходе реакции, выявляют, используя различные методы детекции. Собственно идентификационный анализ ПДАФ хромосомной ДНК в целом аналогичен монолокусному анализу ПДРФ: полиморфные по длине фрагменты, представляющие (как и в случае рестриктазных фрагментов в анализе ПДРФ) разные аллельные варианты гипервариабельных локусов, выступают в качестве индивидуализирующих личность признаков.
Анализ полиморфизма нуклеотидных последовательностей ДНК. Мы уже отмечали, что существенная часть вариаций в полинуклеотидных цепях геномной ДНК обусловлена точковыми нуклеотидными заменами: очень многие локусы, в том числе и высокополиморфные, имеют варианты (аллели), которые на молекулярном уровне одинаковы по длине, но в той или иной степени различаются последовательностью нуклеотидов. Это так называемый сайт-полиморфизм (от англ. «site» - участок), который условно можно обозначить ППАФ - полиморфизм последовательности амплифицированных фрагментов ДНК. Понятно, что амплификация локусов ДНК, обладающих свойством сайт-полиморфизма, приведет к образованию неразличимых по длине фрагментов. Дифференцировать такие фрагменты с помощью обычных методов электрофореза невозможно, поэтому в индивидуализирующих системах ППАФ-типа используются иные принципы дифференциации аллельных вариантов.
Наиболее радикальный путь - секвенирование амплифицированных фрагментов ДНК, то есть определение первичной структуры полинуклеотидной цепи. Теоретически это самый точный и доказательный метод анализа индивидуальных генетических вариаций, поскольку по своей сути секвенирование означает расшифровку генетического кода, который в принципе уникален для каждого организма. В настоящее время единственной молекулярно-генетической индивидуализирующей системой, основанной на секвенировании сайт-полиморфных амплифицированных фрагментов ДНК, являются полиморфные локусы митохондриальной ДНК (мтДНК).
Митохондриальная ДНК человека (см.1.4) представляет собой кольцевую молекулу, которая имеет размер 16 569 пар нуклеотидов и насчитывает всего 37 генов. Она характеризуется рядом уникальных биологических свойств: быстрым темпом мутирования и, как следствие, высоким уровнем изменчивости, большим числом копий в каждой клетке, материнским характером наследования и отсутствием рекомбинации. Эти свойства, ставшие причиной широкого использования мтДНК в популяционных и эволюционных исследованиях, делают ее высокоинформативным, а в некоторых случаях единственно применимым инструментом и в судебно-медицинской практике.
Можно использовать мтДНК как генетический маркер материнской линии наследования, и особенно успешно при установлении родства в тех случаях, когда генетическая дистанция, разделяющая родственников, больше, чем одно поколение. Здесь аутосомные маркеры не дают доказательного результата, поскольку практически невозможно проанализировать очень большое количество вариантов их наследования в ряду поколений. Нуклеотидная же последовательность мтДНК идентична у всех родственников, связанных материнской линией родства (за исключением редких мутационных событий). По этой причине для установления принадлежности индивида к конкретной генетической линии, а, следовательно, для его косвенной идентификации, можно использовать сравнительный анализ мтДНК любых родственников индивида по материнской линии (сестра, бабка, дядя, племянник и т.д.).
Анализ мтДНК возможен, если для экспертного исследования доступны крайне малые количества биологического материала или ДНК, содержащаяся в образце, сильно деградирована и хромосомная ДНК не может быть амплифицирована. Митохондриальная ДНК многократно использовалась для анализа костных останков, возраст которых исчисляется десятками, сотнями и даже десятками тысяч лет. Такие результаты недостижимы для индивидуализирующих систем, основанных на анализе однокопийных генов хромосомной ДНК.
Высокий уровень изменчивости мтДНК обеспечивает широкий спектр индивидуализирующих характеристик, а значит, и высокий дискриминирующий потенциал данного метода. Поскольку мтДНК не подвержена рекомбинации, она должна рассматриваться как единый локус. Количество вариантов последовательности контрольного региона мтДНК существенно превышает количество аллелей любого, даже самого полиморфного хромосомного маркера. Полиморфизм мтДНК столь высок, что иногда один лишь его анализ может обеспечить достаточно надежное подтверждение родства.
Вариации в полинуклеотидных цепях мтДНК обусловлены в основном точковыми нуклеотидными заменами - типичный полиморфизм последовательности ДНК. При типировании мтДНК в судебно-экспертных идентификационных исследованиях используются анализ полиморфизма длины рестриктазных и амплифицированных фрагментов (ПДРФ, ПДАФ), анализ полиморфизма нуклеотидных последовательностей (ППАФ, SSO, гибридизационные микроматрицы) и анализ конформационного полиморфизма (SSCP, DGGE). Тем не менее, следует отметить, что из них практическое значение имеет только индивидуализирующая система ППАФ-типа, в основе которой лежит секвенирование амплифицированных фрагментов ДНК, синтезированных на матрице мтДНК. Такой анализ, предусматривающий прямое секвенирование амплифицированного продукта, дает полную информацию о первичной структуре исследуемого фрагмента (рис.54).
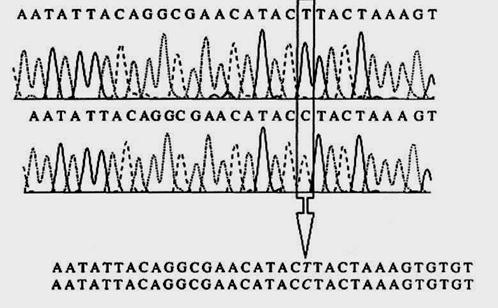
Рис.54. Выявление нуклеотидных замен в митохондриальной ДНК методом прямого флуоресцентного секвенирования
Мы уже говорили, что применительно к задачам судебно-медицинской экспертизы молекулярно-генетический идентификационный анализ наиболее эффективен в двух случаях: идентификация личности и установление биологического родства (в наиболее распространенном варианте - экспертиза спорного отцовства). Под идентификацией понимается, во-первых, задача экспертизы по конкретному уголовному или гражданскому делу, и во-вторых, собственно процесс и результат экспертизы, который заключается в установлении тождества исследуемых объектов (а в случае отрицательного результата - их дифференциации). Предпосылкой для идентификации объекта является его индивидуализация: процесс выявления и оценки признаков объекта, обладающих максимальной значимостью с точки зрения его неповторимости, то есть отличия от иных объектов. Отождествление или дифференциация осуществляются на основании сравнительного анализа совокупности индивидуализирующих признаков ДНК, выявленных в ходе идентификационного исследования.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 1346; Нарушение авторских прав?; Мы поможем в написании вашей работы!