КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Дифференцировка тимоцитов
|
|
|
|
Тимус
ОРГАНЫ ИММУННОЙ ЗАЩИТЫ
Различают центральные (первичные) и периферические (вторичные) органы иммунной защиты. Центральные лимфоидные органы (костный мозг, тимус) — главное место лимфопоэза. Здесь лимфоциты дифференцируются из клеток–предшественниц, размножаются и созревают. T–клетки созревают в тимусе, а B–лимфоциты — в печени плода и костном мозге взрослого организма. В ходе дифференцировки в центральных органах лимфоциты начинают экспрессировать рецепторы, которые в дальнейшем могут связываться с антигеном. В центральных органах отбираются и выживают те лимфоциты, которые толерантны (невосприимчивы) к собственным антигенам. К периферическим (вторичным) лимфоидным органам относят селезёнку, лимфатические узлы, лимфоидную ткань, связанную со слизистой оболочкой (лимфатические фолликулы, миндалины). В периферических лимфоидных органах лимфоциты взаимодействуют между собой, со вспомогательными клетками и с антигенами. Здесь макрофаги, антигенпредставляющие клетки и зрелые Т- и B–лимфоциты участвуют в иммунном ответе, образуются эффекторные клетки и клетки памяти. Иммунные реакции с участием циркулирующих в крови антигенов протекают в селезёнке. Клетки лимфатических узлов реагируют с антигеном, циркулирующим в лимфе. Лимфоидная ткань, связанная со слизистой оболочкой, реагирует на антиген, проникающий в неё из внешней среды.
В тимусе (вилочковой, зобной железе) происходит антиген-независимая дифференцировка T–лимфоцитов. Лимфоциты тимуса называют тимоцитами. Тимус — двудольчатый орган, расположенный в грудной полости над сердцем и магистральными кровеносными сосудами. Каждая доля состоит из долек, которые разделены соединительнотканными трабекулами (рис. 11-10А). Тимус подвергается возрастной инволюции. Атрофия начинается в пубертатном периоде и продолжается в течение всей жизни. Вначале инволюция прослеживается в корковом слое дольки, который может полностью исчезать. Атрофия коркового слоя зависит от чувствительности корковых тимоцитов к стероидным гормонам. Все процессы, приводящие к резкому увеличению уровня стероидов в организме (беременность, стресс), ускоряют атрофию тимуса. Образование T–клеток в тимусе продолжается во взрослом организме, но с меньшей интенсивностью.
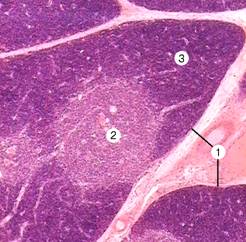
Рис. 11-10А. Вилочковая железа. Прослойки соединительной ткани (септы) делят железу на дольки (1). Более светлая центральная часть дольки — мозговой слой (2), тёмноокрашенная периферическая часть — корковый слой (3). Плотная окраска коркового слоя объясняется большим количеством в нём тимоцитов. Окраска гематоксилином и эозином.
Клетка–предшественница T–лимфоцитов поступает в тимус из костного мозга в плодном периоде (рис. 11-11). Она может экспрессировать на своей поверхности дифференцировочную молекулу CD7. Далее она синтезирует цитоплазматическую форму молекулы CD3, а позже выставляет на поверхности CD1 и CD2.
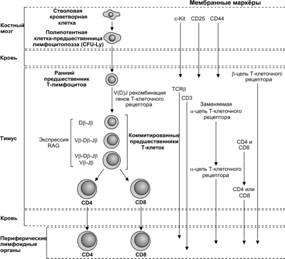
Рис. 11-11. Дифференцировка T–лимфоцитов от стволовой кроветворной клетки до состояния терминальной дифференцировки. [98]
Протимоциты имеют фенотип CD7+CD1+CD2+cCD3+CD4–CD8–. Молекула CD1 исчезает, когда клетка становится зрелой, а цитоплазматическая форма CD3 (cCD3) переходит в мембранную.
Тимоциты. По мере сборки a- и b-цепей рецептора T–лимфоцита протимоциты начинают экспрессировать маркёры CD4 и CD8, давая начало большинству тимоцитов с фенотипом CD1+CD2+CD4+CD8+. Эти клетки способны дифференцироваться в двух направлениях: в клетки CD1–CD2+CD4+CD8- и клетки CD1-CD2+CD4–CD8+ при наличии у обоих подтипов мембранного маркёра CD3 и ab-рецептора T–лимфоцита (рис. 11-12). Этим клеткам разрешено покидать тимус, они появляются в периферической крови и лимфоидных органах. В нормальных условиях выселившиеся из тимуса T–лимфоциты экспрессируют либо CD4, либо CD8, а клетки фенотипа CD4+CD8+ отсутствуют.

Рис. 11-12. Пути дифференцировки T–лимфоцитов. [100]
У ранних предшественников, экспрессирующих CD2 (CD2+CD3–), ещё не произошла рекомбинация генов T–клеточного рецептора и они пока не экспрессируют маркёр T–хелперов молекулу CD4 и общий маркёр для цитотоксических Т-лимфоцитов и Т-супрессоров молекулу CD8. Из всех клеток с фенотипом CD4–CD8– 20% клеток образуют клетки с gd T–клеточным рецептором, 20% с ab T–клеточным рецептором высокой степени гомогенности и 60% клеток, коммитированных к образованию большинства зрелых ab T–лимфоцитов. На последующем этапе дифференцировки CD4-CD8– клетки начинают экспрессировать молекулу адгезии CD44, а затем a цепь рецептора ИЛ2 (CD25). На этой стадии активируется процесс рекомбинации b цепи T–клеточного рецептора, который начинается в группе сцепления D–J генов, а затем в V-DJ. Частота успешной рекомбинацииb цепи возрастает в присутствии двух кластеров гена DJCb. При недостаточной рекомбинации в первом кластере она может быть осуществлена во втором. Вслед за продуктивной рекомбинацией b цепи T–клеточного рецептора в плазмолемму клетки–предшественницы встраиваются молекулы CD3 и заменяемые (суррогатные) a цепи (pTa) по аналогии с заменяемыми (суррогатными) лёгкими цепями в составе предшественника B–клеточного рецептора. Сигнал, действующий через pTa, останавливает рекомбинацию b цепи T–клеточного рецептора, важен для пролиферации клеток и последующей экспрессии CD4 и CD8. На этой стадии из плазмолеммы исчезает CD25. CD4+CD8+ клетки реэкспрессируют белок-активатор рекомбинации 1 и 2 [V(D)J recombination-activating protein 1, или RAG-1 и V(D)J recombination-activating protein 2, или RAG-2] для V(D)J [ V ariable, изменчивый; D iversity, разнообразие; J oining, объединение (генных сегментов)] рекомбинации a цепи, которая может протекать в обеих хромосомах и продолжаться до момента селекции или гибели T–клетки. CD4+CD8+ клетки с ab T-клеточным рецептором мигрируют к границе между корковым и мозговым веществом, где включаются в процесс позитивной или негативной селекции и созревают, превращаясь в CD4+- и CD8+-лимфоциты.
У детей с врождённым отсутствием тимуса вследствие нарушения развития третьего глоточного кармана имеет место дефицит T–лимфоцитов (синдром ДиДжорджи).
gd T–клетки
Механизмы дифференцировки Т- и B–лимфоцитов во многом сходны. Принципиальное различие состоит в том, что предшественники B–клеток вначале экспрессируют IgM, затем одновременно IgM и IgD, а позже один из IgG, IgA или IgE (все с одной и той же лёгкой цепью). В отличие от B–клеток, T–лимфоциты в течение всего жизненного цикла экспрессируют один и тот же T–клеточный рецептор, 95% T–клеток — ab и 5% gd. Рекомбинация генов цепей g, d и b, в которой участвуют молекулы RAG-1, RAG-2 и TdT, протекает практически одновременно и предшествует рекомбинации гена цепи a. Сегменты гена d расположены вблизи области сегментов гена a. Согласно существующему предположению, гены g и d участвуют в рекомбинации первыми и клетка приобретает gd T–клеточный рецептор. Если же рекомбинация генов b цепи опережает рекомбинацию генов других цепей и одновременно с b цепями в плазмолемму встраиваются суррогатные a цепи (pTa), то в подобном предшественнике T–клетки обычно продолжается процесс рекомбинации генов a цепи и в итоге клетка приобретает ab T–клеточный рецептор.
Природа gd T–клеток остаётся неясной. Некоторые из них не рестригированы по MHC, ограничены по идиотипам T–клеточного рецептора и могут формироваться в отсутствие тимуса. Подобные клетки первой волны могут мигрировать в кожу и формировать дендритные клетки. Эти клетки последующей волны заселяют эпителий репродуктивного тракта. Они в небольшом количестве заселяют также периферические лимфоидные органы. Постулируется специфическое взаимодействие gd T–клеток с молекулами повреждённых клеток, таких как белки теплового шока.
|
|
|
|
|
Дата добавления: 2014-12-23; Просмотров: 2861; Нарушение авторских прав?; Мы поможем в написании вашей работы!