КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Формы борьбы за существование 3 страница
|
|
|
|
Если все не слишком вредные, т. е. условно «нейтральные», мутации в процессе дальнейшего мутирования и отбора малых мутаций («модификаторов») теряют свое выражение (в гетерозиготе), т. е. становятся вполне рецессивными, то фенотип не меняется, несмотря на наличие мутаций. Это означает развитие рецессивности мутации и доминантности нормы (в процессе эволюции) и является результатом стабилизирующего отбора.
б. Стабилизирующий отбор в разнородных локально или сезонно меняющихся условиях. Эволюция организмов, живущих в меняющихся условиях разнообразной внешней среды, не Ограничивается выработкой одной ограниченной нормы. В наиболее часто встречающихся типичных условиях преимущества в борьбе за существование будут на стороне некоторой средней нормы, Однако в других реально встречающихся условиях преимущества будут на стороне тех или иных уклонений от этой главной нормы. Точно так же и в различные сезоны требования, предъявляемые к организму, закономерно меняются. Изменяются и направления естественного отбора, и это приводит к установлению «второстепенных норм», т. е. известного полиморфизма (экологического или сезонного), который может иметь генетический характер (гетероморфизм), но может быть и модификационным (собственно полиморфизм). Практически чаще всего происходит комбинирование известного гетероморфизма с выработкой наиболее адекватных форм реагирования. Некоторая зависимость процессов индивидуального развития от изменений во внешних факторах, т. е. известная лабильность, означает и неустойчивость и вместе с тем ненадежность формообразования.
В процессе эволюции наиболее существенные «адаптивные нормы» в известной мере стабилизируются. Это связано с развитием регуляторных механизмов, защищающих эти целостные приспособительные реакции от возможных нарушений со стороны случайных внешних влияний. И это можно рассматривать лишь как результат элиминации всех уклонений от установившихся «адаптивных норм», т. е. типичных форм приспособления к реально встречающимся частным условиям существования. Зависимые процессы развития принимают при этом авторегуляторный характер.
Таким образом, развитие «теневых» и «световых» форм листьев иногда индуцируется заранее внешним фактором. У некоторых деревьев оно определяется условиями освещения в почках, заложившихся в прошлом сезоне вегетации (Nordhausen, цит. по Е. Ungerer, 1926), а затем полностью регулируется внутренним механизмом развития. В этом можно видеть защиту приспособительного формообразования организма от «ошибочных» реакций на переменчивые условия весеннего освещения, т. е. известные черты автономизации развития, которые объясняются только естественным отбором результатов «удачных» реакций, или, иначе говоря, элиминацией всех неблагоприятных уклонений (т. е. «ошибочных» модификаций).
У амфибиотических растений вырабатываются две-три адаптивные нормы для жизни в воде, на болоте или на суше. Они имеют вполне целостный характер и осуществляются при посредстве внутреннего авторегуляторного механизма развития, хотя частично или сполна детерминируются известными интенсивностями внешних факторов. При этом водные листья индуцируются вовсе не обязательно водной средой. У водяной гречихи (Polygonum amphibium) можно получить типичные «плавающие» листья во влажном воздухе, а не только в воде. У стрелолиста (Sagittaria sagittifolia) типичные лентовидные водяные листья развиваются и в воздухе при ослабленном освещении (Vischer, цит. по К. Goebel, 1928). Таким образом, детерминирующим фактором развития здесь является не тот фактор среды, который вызвал к жизни данную модификацию. В процессе эволюции выработался новый механизм индивидуального развития, что, очевидно, возможно только через естественный отбор. Так как отбор происходил на фоне адаптивной модификации, то он, во всяком случае частично, проявлялся в виде стабилизирующего отбора. В данных случаях известная степень влажности и затененности была, очевидно, более надежным раздражителем, чем переменчивый уровень воды в заболоченной местности. Смена детерминирующих факторов должна была произойти в результате элиминации всех неудачных реакций (т. е. «ошибочных» адаптации, приводивших к развитию водной формы на суше, или наоборот).
Подобная стабилизация адаптивных норм наблюдается и у животных, однако, главным образом, в качестве сезонных форм (подвижность большинства животных не ставит перед ними вопроса о локальных экологических адаптациях). Вполне стабильными являются сезонные формы бабочки Araschnia levana-prorsa. Довольно стабильны также, и во всяком случае вполне характерны для различных видов, и сезонные формы коловраток и дафний. Все они определяются в своей реализации не столько внешними факторами, сколько существованием определенных систем внутренних регуляторных механизмов. Авторегуляторный характер развития сезонных форм совершенно ясен. Нужно думать, что и здесь регуляторные механизмы развились в результате действия стабилизирующего отбора при устранении всех уклонений от приспособленной в данных условиях частной (сезонной) нормы.
в. Стабилизирующий отбор в колеблющихся условиях внешней среды. В любых условиях существования внешние факторы подвержены более или менее значительным случайным и кратковременным изменениям (нерегулярным колебаниям интенсивности). Однако в тех случаях, когда эти колебания очень резко выражены, стабилизация формообразования приобретает исключительно большое значение. Лабильные организмы реагируют на эти изменения, однако новые изменения в факторах среды лишают эти реакции их приспособительного значения. Реакция оказывается «преждевременной» и потому «ошибочной». У лабидьных организмов подавляющая масса неблагоприятных уклонений составляется из таких «ошибочных» модификаций, и потому интенсивная их элиминация должна вести к максимальной стабилизации форм. Стабилизирующий отбор должен идти с большей скоростью.
Возможность преобразования лабильного признака в стабильный показана экспериментально М. Камшиловым путем отбора на максимальное выражение при противоположных изменениях условий развития. Искусственным отбором удалось достигнуть того, что мутация eyeless выражалась одинаково как при воспитании на влажном корме (нормальный максимум), так и при воспитании на сухом корме (нормальный минимум). Получена экспериментальная автономизация развития данного признака, превратившегося из зависимого в независимый от изменений в факторах внешней среды (М. Камшилов, 1939в).
В природных условиях наиболее значительны колебания температуры, влажности и инсоляции в горных местностях, в резко континентальных и, наконец, в полярных странах.
В этих условиях действительно происходит иногда весьма быстрая стабилизация новых форм.
Н. И. Калабухов и В. М. Родионов (1936) показали, что равнинные предкавказские лесные мыши при перенесении в горы дают типичную горную реакцию в виде увеличения числа эритроцитов и количества гемоглобина в крови. Однако горные формы того же подвида Apodemus sylvaticus ciscaucasicus при перенесении на равнину уже не дают обратной реакции. Повышенное содержание гемоглобина у них наследственно стабилизировалось.
Обыкновенная пастушья сумка (Capsella bursa pastoris) имеет в горах Эрдшиас-даг в Малой Азии типичный альпийский характер — глубокие корни, низкий стебель, ксерофитный габитус с опушением. Хотя это растение, по Цедербауеру (Zederbauer, 1908, цит. по Weidenreich, 1921), могло быть занесено сюда только человеком и, следовательно, лишь недавно стало альпийским растением, оно при высеве семян в низменности в течение более четырех поколений все же сохраняло низкий стебель (корни и листья теряли альпийский характер). Следовательно, процессы формирования стебля здесь уже полностью стабилизировались (автономизировались).
Таких фактов частичного наследственного «фиксирования» изменений, которые ранее были зависимыми, т. е. имели характер приспособительных модификаций, известно очень много. Таковы различные экологические и географические формы как растений, так и животных, которые при переносе в иные условия, близкие к исходным, дают неполный возврат к прежнему фенотипу. Возможность возврата указывает на значение модификации в возникновении данной формы. То обстоятельство, что этот возврат является лишь частичным (или в известных признаках), показывает, что данный фенотип был все же объектом естественного отбора (и именно стабилизирующего, так как он происходил в пределах установившейся уже модифицированной нормы). Длинная и тонкая шерсть полярных млекопитающих (также, короткие уши и хвосты) есть наследственное их свойство. Однако при выращивании в более теплом климате эти признаки частично изменяются. С другой стороны, и млекопитающие умеренных стран получают в более холодном климате более длинную шерсть. Это — модификации, на фоне которых произошла стабилизация процессов развития более мощного шерстного покрова полярных животных (путем элиминации всех особей, которые в результате реакции на случайно более высокую температуру среды развивали недостаточный шерстный покров).
Если альпийский габитус развивается в горах у очень многих растений, то большинство типичных альпийских растений сохраняет хотя бы частично свой альпийский габитус и при переносе в долины. Формообразование их стало более стабильным, очевидно, в результате жесткой элиминации морозами и засухой всех особей, которые в случайно мягкие и влажные весны давали «ошибочную» долинную модификацию (с длинными стеблями, не-опушенными листьями и короткими корнями). Частным и притом более простым случаем стабилизации в условиях резких колебаний в факторах внешней среды является также распространение полиплоидных растений в горных, резко континентальных и полярных странах (Е. В. Вульф, 1937). Полиплоиды отличаются именно более устойчивым формообразованием.
В. СТАБИЛИЗАЦИЯ НОРМЫ В ПРОЦЕССЕ ЭВОЛЮЦИИ
Если мы рассматривали отдельно ведущую и стабилизирующую формы отбора, то это было сделано лишь для внесения максимальной ясности. На самом деле обе формы естественного отбора действуют всегда совместно, так как, с одной стороны, внешняя среда постепенно, но неуклонно меняется, и, следовательно, могут быть обнаружены ведущая роль отбора и наследственное изменение нормы, но, с другой стороны, это — процесс медленный и стабилизирующий отбор всегда и непрерывно ведет к развитию регуляторных механизмов, охраняющих эту медленно меняющуюся норму от нарушающих ее внешних влияний.
Ведущая форма отбора яснее всего устанавливается у стабильных организмов с автономным развитием, так как наследственные изменения у них не осложняются результатами индивидуального приспособления. Стабилизирующая форма отбора устанавливается и у «стабильных» организмов, но протекает более типично у лабильных организмов и особенно в условиях колеблющейся среды (с изменениями случайного характера). Однако именно в этих случаях трудно разграничить результаты ведущей формы отбора, меняющей норму реакций, и индивидуального приспособления, основанного на этих реакциях и дополняемого стабилизирующим отбором. Эта трудность нашла свое отражение и в нашем изложении: фактический материал, использованный в качестве иллюстраций, в сущности один и тот же. В некоторых случаях особенно ясно видно соединение обеих форм отбора в процессе эволюции (т. е. в процессе изменения самой нормы).
Известно, что при засухе листья вянут и затем опадают. В тропических странах нередко наблюдается вполне закономерный листопад в засушливое время года. Этот листопад является приспособлением, предохраняющим растение от потери воды при транспирации через листья и невозможности пополнения ее запаса через корневую систему. В умеренном климате зимой, при замерзании воды, создаются также условия «засухи», и это ведет также к листопаду (G. Nichols, 1924).
При экспериментальном удалении листьев у многих растений (Vicia faba, Sarothamnus scoparius, Lathyrus odoratus, Syringa vulgaris, Rosa centifolia, Robinia pseudoacacia и др.) сильно увеличивается число хлорофилльных зерен в ассимиляционных тканях стеблей и черешках листьев, а также происходит и новообразование хлоропластов в нижележащих слоях клеток. Клетки ассимиляционной ткани удлиняются в радиальном направлении, и возникает палисадная ткань (если ее не было; а если она была, то она сильнее разрастается). Число устьиц значительно возрастает. Экспериментально установлено усиление ассимиляции и транспи-рации (A. Boirivant, 1897, и Braun, 1899; цит. по Ungerer, 1926). Такая компенсация имеет, следовательно, приспособительный характер и приобретает особое значение у растений засушливых местностей, нередко сбрасывающих листья. В этом случае черешки и стебли функционально замещают сброшенные листья.
Сбрасывание листьев при засухе может оставлять растение безлистным на длительный период вегетации. Ксерофитное растение может в молодом возрасте развивать листья, а затем их окончательно терять (многие акации). Некоторые ксерофитные кустарники напоминают тогда безлистную метлу. Таковы некоторые спаржевые, но в особенности многие санталовые и мотыльковые, как, например, ракитник (Cytisus), бобровник (Spartium). Ассимилирующая ткань под кожицей стеблей обладает сильно редуцированными межклетниками. Устьица многочисленны, но малы и погружены. Здесь адаптивная модификация приобрела постоянный характер. В дальнейшем процессе эволюции побеги частью расширяются и образуют листообразные ветки — «филло-кладии» (например, иглицы — Ruscus hypoglossum и Ruscus acw leatus). У других растений расширяются не стебли, а черешки сбрасываемых листьев — «филл одни» (особенно у акаций). У Acacia heterophylla и др. в нормальном онтогенезе развиваются филлодии, и после этого опадают листья (рис. 11). Сбрасывание листьев и развитие филлодиев определяются уже автономно, т. е. внутренними факторами развития, а не засухой и компенсационной модификацией, связанной с отсутствием листьев. Таким образом, в процессе эволюции создается трудно понимаемая замена листьев другими сходными образованиями, несущими ту же функцию. Казалось бы, как это и происходит у других растений, сами листья могли бы приспособиться к условиям засухи.

Рис. 11. Молодое растение Acacia pyienantha. Первичные перистосложные листья (1–4) сменяются переходными листьями (5, 6) и развивающимися позднее типичными филлодиями (7 — 9). По К. Циммерман, 1930
В данном случае этот своеобразный путь эволюции объясним только как результат обусловленного засухой периодического листопада с последующими компенсационными явлениями. Здесь именно адаптивная модификация тканей стеблей и черешков имела ведущее значение в процессе дальнейшей эволюции. Эта модификация приобрела в ксерофитных условиях (очевидно, через механизм стабилизирующего отбора) значение постоянного, т. е. стабильного признака организации, развивающегося и без засухи. В процессе обычного отбора произошло затем приобретение листообразной формы новых органов, более приспособленных к ограниченной транспирации в сухой местности. Эти данные показывают с достаточной убедительностью, что именно вынужденная (периодической засухой и листопадом) адаптивная модификация была переходным этапом на пути эволюции, приведшей к образованию новых ассимилирующих органов (филлокладиев и филлодиев).
Стабилизация происходила, очевидно, на базе элиминации тех особей, которые давали в условиях случайно более влажной весны прежнюю реакцию нормального листообразования и затем, летом, страдали от чрезмерного испарения, пока не опадут листья, и от недостаточной ассимиляции углерода, пока не наступят компенсаторные модификации в стеблях и черешках. Здесь ясна и роль приспособительной модификации в процессе эволюции, и роль стабилизирующей формы отбора, ведущей к установлению внутреннего механизма развития этой модификации, и движущая роль обычного отбора, ведущего к развитию новой нормы с новыми листообразными органами.
Мощное развитие жевательной и височной мышц хищных млекопитающих сопровождалось прогрессивным развитием зубной системы, челюстей, венечного отростка нижней челюсти, скуловой дуги, а также сагиттального гребня черепа. Вряд ли можно сомневаться в том, что ведущим в этом процессе эволюции хищных млекопитающих было изменение инстинктов животного, переходившего от питания насекомыми к добыче и использованию более крупных животных. Первым результатом такого перехода могла быть только усиленная тренировка челюстных мыши с ее последствиями — увеличением их массы (а также их кровоснабжения) и увеличением поверхности их прикрепления на частях скелета. Мы знаем, что такие изменения могут иметь характер приспособительных модификаций (опыты Хюртля) и не лишены целостности. Однако в процессе эволюции несомненно происходила стабилизация таких изменений (через элиминацию особей с недостаточным развитием челюстей и их мускулатуры, случайно выросших в благоприятных условиях питания мелкими животными) на почве индивидуальной конкуренции в добывании крупных животных. Соответственно происходила «фиксация» новой нормы (при этом менялся и способ ее развития — в автономном процессе формирования мышцы ее объем определяется числом волокон, а не их толщиной, как при тренировке). Однако в то же самое время известные преимущества в борьбе за существование имели и те особи, которые обладали наследственно еще более совершенной нормой и, в частности, особи с наследственными изменениями в зубной системе (которая, как известно, от «упражнения» заметно не меняется, во всяком случае форма и величина зубов не модифицируются). Поэтому и ведущая форма отбора имела здесь бесспорное значение.

Рис. 12. Посевы Camelina linicola и С. glabrata, сделанные одновременно и выращенные в совершенно одинаковых условиях. Видно наличие наследственных различий нормы реакций. Вверху — С. linicola, внизу — С. glabrata. По Н. В. Цингеру, 1909
Значение приспособительной модификации, как переходного этапа в процессе эволюции, играющего в течение первоначального периода формирования новой адаптивной нормы (или замены главной нормы добавочной) руководящую роль, ограничивается скоростью этих изменений. Эта скорость так значительна, что при быстрой смене условий внешней среды приобретает характер решительного скачка, сразу переносящего организм в новую среду. Мы уже отмечали скорость этого процессса на примерах горной формы Capsella bursa pastoris. To же относится к модификации ярового рыжика Camelina glabrata, приведшей в посевах льна, т. е. за время развития этой культуры, к образованию льняного рыжика С. linicola. Образование высокого стебля с длинными междоузлиями и узкими листьями, возникшее первоначально как модификация (как это наблюдается при высеве С. glabrata в посевах льна), здесь уже стабилизировалось, т. е. наследственно «зафиксировано». С. linicola дает высокие стебли с узкими листьями и вне посевов льна (рис. 12, 13). С другой стороны, увеличение размера семян представляет результат прямого отбора в его ведущей форме (Н. В. Цингер, 1909).
Конечно, этот быстрый переход возможен лишь на основе, подготовленной предшествующей эволюцией организма. Ведь сама способность организма к адаптивной модификации является частью унаследованной нормы его реакций, выработанной ведущей формой отбора.
Это особенно ясно в случаях резко выраженного полиморфизма, т. е. существования нескольких целостных адаптивных норм. У большинства высших растений поверхность листьев, их строение, опушение, толщина кутикулы и численность устьиц легко подвергаются явным модификациям в зависимости от интенсивности освещения, так что говорят о световых и теневых листьях. В частности, обе реакции хорошо выражены у вербейника (Lys-machia vulgaris). Однако теневые формы Lysimachia numullaria и Dactylis glomerata сохраняли свои характерные признаки и при длительной культуре (многие годы) на солнечном месте (Turesson, 1922). Здесь световая модификация оказалась утерянной, а теневая стабилизировалась.
Иногда и длительная история организма в новой среде не исключает возможности частичного возврата к исходным формам при восстановлении прежних условий. Так, по исследованиям Н. Ва-куленко (1940), при прорастании зимних почек типичного водяного растения Myriophyllum verticillatum в воздушной среде (во влажной камере) на верхних частях стебля развивались еще цельнокрайние листья. Здесь совершенно ясен переход от амфи-биотического растения с двумя нормами (причем первично главной нормой была наземная, а добавочной — водяная) к чисто водяному растению, почти уже утратившему свою прежнюю наземную норму и стабилизировавшему развитие своей новой нормы.

Рис. 13. Сравнение Camelina linicola с различными модификациями С. glabrata. Нормально льняной рыжик растет в густых посевах льна. 5 — С. linicola, выращенный в редком посеве; 6 — С. glabrata, выросший в тех же условиях, что и С. linicola на рис. 5. Различия между 5 и б имеют генотипический характер; 7 — С. glabrata, развивавшийся в посеве льна, соответственно обычным условиям развития С. linicola; 8 — С. glabrata, развившийся на просторе. Оба последних экземпляра получены из семян одной особи. Различия между 7 и 8 покоятся на модификациях. Внизу показаны 4-й, 6-й, 9-й, 12-й и 15-й листья соответствующих растений. По Н. В. Цингеру, 1909
Такой переход реализуется, конечно, с большой легкостью и скоростью при подходящих для этого внешних условиях, когда те факторы, которые раньше имели лишь локальное или временное значение, становятся постоянными для данного вида организмов. Изложенное представлено на рис. 14, А, В, С, Д.
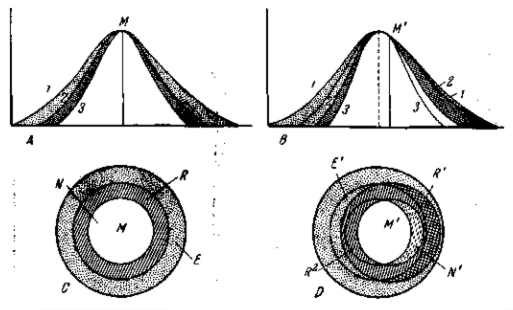
Рис. 14. Отбор в разнообразных и колеблющихся условиях внешней среды (с учетом различных форм индивидуального реагирования, т. е. модификаций). А — отбор при установившихся (в данном многообразии) условиях (стабилизирующая форма отбора). Вариационные кривые: 1 — всех особей; 2 — особей, вышивающих в реально встречающихся различных условиях (условно «нормальные» особи, условно благоприятные и частично благоприятные уклонения); 3 — особей, выживающих и оставляющих потомство в обычной средней «нормальной» обстановке. Точками обозначена зона безусловной элиминации уклонений, вредных при всех условиях (мутаций и морфозов). Косой штриховкой обозначена зона вариантов, элиминируемых при обычных, нормальных условиях, но выживающих в известных уклоняющихся, но реально встречающихся условиях. В — отбор при изменении условий в определенном направлении (с сохранением их многообразия). Ведущая форма отбора. Вариационные кривые: 1 — всех особей; 2— особей, выживающих в различных условиях новой среды; 3 —особей, выживающих в новой «нормальной» обстановке. С, D — другой 'план изображения механизма стабилизирующей и ведущей форм отбора.
Эволюция половых признаков — одна из интереснейших биологических проблем, до настоящего времени еще далеко не разрешенная. Своеобразие этой эволюции обратило на себя внимание Ч. Дарвина, отметившего многие противоречия тем закономерностям, которые были им установлены для общей организации живых существ. Дарвину пришлось ввести особый принцип полового отбора для объяснения части этих противоречий. Однако тот же Дарвин обратил внимание и на то обстоятельство, что многочисленные факты передачи вторичных половых признаков с самца на самку противоречат и этому принципу. Относящиеся сюда факты не получили своего объяснения и до настоящего времени. Я здесь не принимаю во внимание физиологических объяснений передачи самцовых признаков предположениями о повышении концентрации мужского гормона у самки. Если бы они и оказались в известных случаях верными, то, вскрывая механизм этой передачи, они все равно потребовали бы раскрытия причин исторического возникновения этого механизма. Дело касается, однако, не только вторичных половых признаков. И в эволюции копулятивных органов, их вспомогательных частей и даже самих половых желез мы можем отметить немало явлений, непонятных с обычных точек зрения.
Одним из важнейших доводов в пользу эволюционного учения были в свое время факты наличия у эмбрионов или личинок зачаточных органов провизорного значения, которые не достигают полного развития у данного организма, но вполне сравнимы с зачатками дефинитивных органов других организмов. Такие зачаточные органы мы считаем унаследованными от предков, у которых они были вполне развиты. Это — явления так называемой рекапитуляции предковых состояний.
Однако в половой системе мы находим зачатки подобного же характера, которые не могут быть обозначены как рекапитуляция. У самца млекопитающих в течение эмбрионального периода развиваются яйцеводы и зачаточная матка (uterus masculinus), а у самки развивается копулятивный орган, превращающийся позднее в клитор. Считается, что половая система развивается на некоторой «индифферентной» основе, общей для обоих полов. Эта основа, однако, вовсе не индифферентна: она попросту совмещает дифференцированные признаки обоих полов. Млекопитающие не произошли от гермафродитов — их предки были раздельнополыми. Следовательно, в этом совмещении зачатков органов обоего пола нет рекапитуляции. Самцы никогда не обладали яйцеводами и маткой, а самки не обладали копулятивным органом. Их зачатки не являются у них повторением предковых состояний. Наоборот, они возникают сначала у одного пола, затем как эмбриональные признаки у другого пола, и, наконец, переносятся в большей или меньшей мере и на взрослое животное. Можно было бы думать, что это связано с самим механизмом наследования, в котором участвуют оба пола. Однако и это не так просто. Механизм раздельного наследования половых признаков имеется с самого начала их появления — половые признаки возникают именно у одного из полов. Только в дальнейшей эволюции раздельное наследование нередко почему-то нарушается.
Это нарушение, ведущее к передаче признака на другой пол, иногда определяется выгодой и связано с действием естественного отбора (например, передача органа защиты — рогов — с самца на самку или передача покровительственной окраски самки на самца). Такие случаи разобраны Дарвином. Однако они не составляют общего правила. Наоборот, гораздо чаще наблюдается другое: передача признаков с одного пола на другой не связана с ясной выгодой и поэтому вряд ли объяснима действием обычного естественного отбора. Во многих случаях, конечно, трудно судить о биологическом значении даже таких признаков, как рога самки. Не всегда они имеют значение органов защиты. Однако нередко мы не только не видим положительного значения передачи полового признака с одного пола на другой, но можем даже предполагать, что эта передача скорее неблагоприятна. Для вторично-половых признаков это иногда кажется почти бесспорным.
В самом деле, если вторично-половой признак, например самца, имеет известное биологическое значение (привлечение и возбуждение самки, стимуляция нормального полового цикла у самки и, в результате этого, обеспечение спаривания и оплодотворения), то с передачей его на самку он тем самым теряет свое значение у самца, т. е. обесценивается. С другой стороны, самка приобретает ненужные ей органы (например мозоли на задних конечностях у некоторых амфибий, шпоры на ногах у некоторых куриных), теряет свои средства пассивной защиты (покровительственную окраску) и приобретает яркую, бросающуюся в глаза окраску, делающую ее более легкой добычей хищников. Приобретение самкой яркого оперения, как правило, не только биологически бесцельно, но и прямо вредно. Поэтому в процессе эволюции «независимых» половых признаков самца у самки вырабатывается особый тормозящий механизм, противодействующий этой передаче. И все же передача нередко осуществляется вопреки наличию такого механизма.
Нередко такая передача признака с самца на самку имеет резкий, скачкообразный характер — она не подготовляется в процессе медленной эволюции самки. И в этом мы видим своеобразие, нуждающееся в объяснении. И, наконец, соответственно двум разным механизмам индивидуального развития вторично-половых признаков самца — как зависимых от мужского полового гормона или от него независимых (с торможением женским гормоном у самки) — имеются и два разных способа передачи сам-цовых признаков с самца на самку: через повышение чувствительности тканей к мужскому половому гормону или через понижение их чувствительности к женскому гормону (Б. Г. Новиков, 1940). Два разных пути ведут к одному и тому же результату. Очевидно, в основе этих явлений лежат какие-то более сложные биологические закономерности, а не простые физиологические процессы, связанные с механизмом определения пола.
Однако обычное объяснение исторических закономерностей направлением естественного отбора, идущим на основе селекционного преимущества известных уклонений (в данном случае самок, обладающих самцовыми признаками), очевидно, также недостаточно.
1. Передача распознавательных вторично-половых признаков с одного пола на другой. Основные факты, касающиеся явления передачи самцовых признаков самкам, собраны уже у Ч. Дарвина. У оленей, как правило, рога имеют только самцы. Только у северного оленя самка обладает хорошо развитыми рогами. Однако Дарвин отмечает, что у самок некоторых видов оленевых постоянно или случайно встречаются следы рогов. Например, большинство самок вапити (Cervus canadensis) на месте рогов имеет острый костяной выступ. У антилоп имеется полный ряд форм — от видов с рогами только у самцов, через такие виды, у которых самки имеют ничтожные зачатки рогов или очень небольшие рога, до видов с самцами и самками, несущими одинаковые рога. У некоторых животных рога имеют обычно и самцы, и самки, однако у самцов они, как правило, значительно больших размеров (козлы, бараны, быки). При кастрации самца его рога получают особенности рогов самки. Ч. Дарвин отмечает вообще различное влияние кастрации самца на развитие рогов у оленей и у полорогих жвачных.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 560; Нарушение авторских прав?; Мы поможем в написании вашей работы!