КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Формы борьбы за существование 7 страница
|
|
|
|
Африканские сомы рода Clarias образуют непрерывный ряд видов, ведущий от Clarias lazera Си v. et Val. с широкой лентой сошниковых зубов в Сев. Африке через С. mossambicus (Peters) из области озер Виктория и Танганьика и С. gariepinus (Burchell), распространенной от Замбези до Оранжевой реки, и до С. сареп-sis С. u v. et V а 1. с узкой прерванной полосой сошниковых зубов, живущей только в Натале (Worthington, 1940). Однако естественно, что на этих животных наиболее ясно видны результаты территориального разграничения у сидячих и малоподвижных форм. Поэтому классические примеры территориальной дифференциации относятся к наземным моллюскам (исследования Гулика, 1905, над улитками рода Achatinella на Гавайских островах, исследования Крэмптона, 1919, 1925, 1933, над улитками рода Partula на островах Таити и Муреа, исследования Плате, 1907, над улитками рода Cerion на Багамских островах), а частью и к донным, прибрежным формам (Purpura lapillus у Британских островов, по Куку). У морских моллюсков малая подвижность большинства форм компенсируется некоторой подвижностью личинок (в планктоне). Однако и планктонные организмы не рассеиваются особенно широко, даже когда они переносятся широкими массами в морских течениях, так как сохраняют пространственную связь между родственными особями. Даже активно плавающие и далеко мигрирующие организмы нередко держатся совместно, плавая целыми стайками. Образование многих элементарных экологических рас или линий у обыкновенной сельди (по Гейнке, Heincke, 1898) объясняется именно их инстинктами, благодаря которым целые выводки держатся в своих миграциях совместно и возвращаются для икрометания на прежние места. Это и поддерживает у них родственное скрещивание, дающее возможность закрепления вновь возникающих удачных комбинаций. То же самое касается, по-видимому, и различных лососевых и ряда других мигрирующих рыб. Тем более это относится к рыбам с оседлым образом жизни, подобным береговой бельдюге (Zoarces viviparus), которая образует множество местных рас, живущих в отдельных фиордах Скандинавии (Schmidt, 1917). Однако то же самое встречается и у таких подвижных животных, как птицы, в том числе перелетные. Обыкновенный аист (Ciconia ciconia), гнездящийся в Европе и зимующий в Африке, возвращается ежегодно IK одному и тому же гнезду, а молодь возвращается к местам своего вывода и гнездится в ближайшем соседстве. Фактически скрещивание происходит в очень тесной популяции родственных форм и лишь изредка освежается случайным прибытием мигранта.
|
|
|
То же самое касается ласточек. Меченая молодь возвращалась после зимовки на места своего рождения и гнездилась в пределах двух километров от материнского гнезда (Wachs, 1926). Ренш (Rensch, 1939) сопоставил результаты видовой дифференциации у больших птиц с широким распространением и миграцией, у малых перелетных птиц и у малых оседлых (по сводке птиц Пале-арктики Хартера). Из этого сопоставления ясно видно значение оседлости для дифференциации видов. Очевидно, родственное скрещивание (т. е. ограничение панмиксии) способствует выделению новых форм.
Интересный пример значения оседлости для самого начального разграничения популяций дали наблюдения Промптова над зябликом, который в южных областях СССР распадается на ряд популяций, ограниченных известным ареалом обитания и различающихся только по пению. Весьма вероятно, что эти различия не имеют еще ясно выраженной наследственной базы — молодь учится петь у родителей и других взрослых птиц ближайшего соседства. Такое обособление возможно, однако, лишь при условии возвращения после перелета в прежние места обитания (Промптов, 1930). В Московской области зяблик разбился на две экологические формы, различающиеся только по выбору мест для гнездостроения: одна гнездится в лесу таежного типа, смешанном со значительной примесью ели, а другая — в невысоких и редких, с полянками, молодых еловых и??? рощах (Промптов, 1934)
|
|
|

Если это касается таких подвижных животных, как рыбы, с одной стороны, и птицы — с другой, то в еще большей мере то же применимо и к любым другим животным. Экспериментальные исследования Нобля (Noble, 1934) над ящерицей Sceleropus undulatus показали это с полной ясностью. Этот автор следил за переменой мест обитания ящериц, помеченных летом на их естественном местопребывании. Из 226 меченых ящериц к следующему лету сохранилось на прежнем месте 155. Многих из них видели многократно. Наибольшее отдаление местонахождений от исходного места достигало лишь 250 футов. При искусственном перенесении на более значительное расстояние ящерицы возвращались назад.
У автора создалось впечатление, что каждая особь охотится в пределах своей, очень ограниченной территории, лишь изредка заходя на территорию соседа.
Как известно, то же самое касается многих хищных птиц, а также и млекопитающих, держащих под своим контролем определенную территорию, необходимую для их прокормления. В особенности это относится ко многим грызунам, в том числе и крупным (бобры), а также к крупным хищникам.
Еще большее значение, чем обычная оседлость, имеет, конечно, сидячий образ жизни. Из животных он свойствен исключительно водным, так как только в воде может быть обеспечен пассивный перенос пищевого материала. С другой стороны, для растений только в воде возможна известная подвижность молодого или зрелого организма.
Прикрепленный образ жизни ведет, в общем, к тем же результатам, как и оседлость. Во всех этих случаях имеются средства для перемещения и распространения. Сидячие животные обладают подвижными личинками, а у растений пыльца переносится ветром или насекомыми и семена рассыпаются и подхватываются также ветром или высшими животными. Таким образом осуществляется панмиксия, а следовательно, и непрерывное перекомбинирование наследственного материала. Однако практически эта панмиксия значительно ограничена прикрепленным образом жизни. Подвижность личинок сидячих животных невелика, и они в большинстве прикрепляются поэтому недалеко от родительских особей, так что и скрещивание фактически ограничивается главным образом близкими родственниками. Да кроме того, сидячие животные часто обладают еще способностью к вегетативному размножению, т. е. к образованию генетически вполне однородных колоний. У растений, хотя пыльца и разносится, все же наиболее велики шансы опыления цветов собственной пыльцой, пыльцой с других цветов той же особи или от соседней особи. Семена же практически высыпаются в ближайшем окружении родительской особи. Образование родственных колоний поддерживается нередко способностью к вегетативному размножению.
|
|
|
Наконец, к тому же ведет и то обстоятельство, что почти никогда ни растение, ни животное не встречает необходимых для него экологических условий на больших сплошных пространствах. Обычно, даже при сплошном географическом распределении, подходящие экологические условия (почвы, их влажность, освещение, сообщество других организмов) встречаются лишь пятнами, которые тогда и оказываются средоточием колонии близко родственных особей. Величина таких колоний иногда ничтожна и при условии полной изоляции они были бы обречены на гибель, в силу утраты пластичности. Так как, однако, обмен мигрантами все же время от времени осуществляется, то и генетический состав популяций оказывается подвижным. Вместе с тем в каждой популяции фиксируются свои особенности, свои комбинации, которые испытываются в борьбе за существование и могут быть исходными для более широкого распространения.
В самом деле, как показали исследования Андерсона (Е. Anderson, 1936) над колониями Iris virginica, такие генетические различия между отдельными колониями вполне уловимы.
2. Экологическая изоляция. Экологическая изоляция, как упомянуто, не всегда отграничима от территориальной и географической, поскольку географические различия могут сопровождаться экологическими, а экологические формы разделяются и территориально. В особенности это касается растений, у которых, однако, иногда особенно ясно выступает именно экологическое расчленение как начальный этап образования новых форм. Таково значение почвенных условий и условий освещения в формировании экологических рас (экотипов), которые, возникая как экологические формы, оказываются сразу расчлененными и территориально, что способствует родственному скрещиванию внутри каждой новой группировки. Поскольку вегетация и время созревания у растений весьма зависимы от экологических условий, такое обособление может непосредственно привести и к различиям во времени созревания, и тогда экологическая изоляция дополняется биологическим ограничением возможности скрещивания с исходной формой. Так, по Андерсону (Е. Anderson), обособление близких видов Iris обусловлено в значительной мере расхождением во времени цветения. У животных некоторое расхождение в местообитании, в пищевом режиме, в образе жизни (дневной или ночной) связано с уменьшением шансов на встречу различающихся в этих отношениях особей, а следовательно, также способствует скрещиванию внутри обособливающихся групп. В основе этого расхождения лежит у животных, очевидно, некоторое изменение инстинктов, ведущее к изменению образа жизни, питания и т. п. Поэтому зоологи рассматривают иногда экологическую изоляцию как частную форму физиологической. Типичным примером начального обособления экотипов у животных могут служить песчаная и солончаковая формы муравья Cardiocondyla stambulovi For. (К. В. Арнольди, 1939).
|
|
|
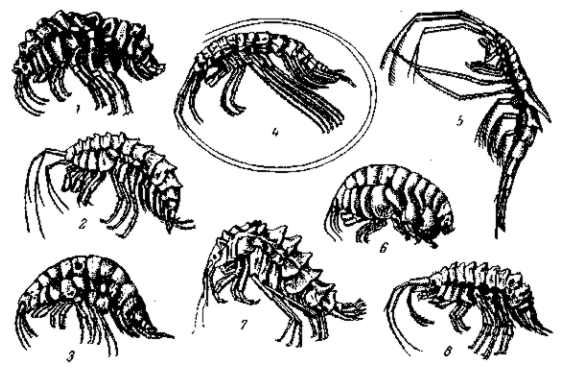
Рис. 17. Некоторые байкальские бокоплавы. 1 — Brandtia lata lata (Dyb.); 2— Parapallasea lagowshii (Dyb.); 2 — Ommatogammarus albinus (Dyb.); 4 — Garjajcwia sarsi (Sow.); 5 — Marcrochestopus branichii (Dyb.); 6 —Gammarasphaera insularis (Bas.); 7 — Acanthogammarus victorii (Dyb.); 8 — Spinacanthus parasiticus (Dyb.). По А. Я. Базикаловой, 1940
Типично экологический характер имеет процесс обособления нескольких рас мышей Peromyscus maniculatus, живущих в одном и том же ареале, но предпочитающих в одном случае лес, в другом случае — берега озер (Dice, 1931) или открытые места. Различия в окраске некоторых видов грызунов Perognathus связаны с различиями в цвете почв их местообитаний (Benson, 1933). Эти различия имеют экологический характер. Они связаны, однако, и с пространственным разграничением. Экологический характер имеет и распределение горных растений и животных на разной высоте. Множество эндемичных видов бокоплавов (gammaridae) оз. Байкал должно было образоваться на месте, на базе экологического расхождения немногих родоначальных форм по различным грунтам, зарослям, по разным глубинам и т. п. (рис. 17). Сезонное разделение произошло у рас обыкновенной сельди, которые мечут икру либо весной в различных местах прибрежного мелководья, либо осенью на мелях более открытого моря. Сезонные расы, отличающиеся по месту и времени икрометания, известны и для целого ряда проходных и полупроходных пресноводных рыб из семейств лососевых, осетровых, карповых, а также у судака (Л. Берг, 1934). Обыкновенная креветка (Crangon vulgaris) известна в двух расах, еще не различимых по виду, из которых одна мечет икру весной, другая — осенью (L. Plate, 1908). Дубовый шелкопряд (Lasiocampa quercus) появляется в июле — августе, а его почти неотличимый подвид (L. guercus callunae) — в мае — июне, так что скрещивание между ними почти невозможно. Бабочки Agriodes thetis и A. coridon различаются тем, что второй вид имеет один период размножения, падающий между двумя периодами размножения первого. Близкие виды бабочек — Eupithecia innotata Hufn. (выводится на Artemisia) и Е. unedo-nata Mob., которая выводится много раньше (на Arbutus), нормально не могут скрещиваться. Однако путем охлаждения куколок второго вида можно задержать вылупление, и тогда удается произвести скрещивание и получить вполне плодовитые гибриды Dietze. Такие же различия имеются между расами многих других бабочек (многие шелкопряды, в том числе тутовый), образующих различное число поколений в течение одного сезона. Фактически не скрещиваются в силу расхождения во времени вылупления и две мало различимые расы 17-летней цикады (Tibicen septemdecem L).
Во всех этих случаях едва намечающиеся различия сопровождаются значительной затрудненностью скрещивания между расходящимися формами вследствие разновременности созревания половых продуктов. Это — безусловно решающий момент, обеспечивающий фиксирование новых форм и их расхождение.
3. Физиологическая изоляция. Под физиологической изоляцией понимают затрудненность или малую эффективность скрещивания между представителями разных групп особей (несмотря на полную возможность их встречи).
В основе физиологической изоляции лежит как генотипическое и биохимическое, так и физиологическое и морфологическое расхождение признаков. В качестве последнего можно отметить расхождение в строении полового аппарта, которое делает копуляцию с исходной формой невозможной. Несомненно, такое расхождение иногда предшествует обособлению по другим морфофизио-логическим признакам (часто у насекомых). Однако оно все же представляет уже результат эволюции, связанной с взаимным приспособлением полов. Такое расхождение не может быть результатом единичной мутации (за исключением различий в общей величине тела, которые также иногда ведут к невозможности скрещивания), так как взаимность приспособления требует сложного согласования ряда отдельных изменений у обоих полов. То же самое касается видовых отметин и вторично-половых различий, которые должны быть также согласованы с изменениями в инстинктах и рефлексах, ведущих к спариванию. И такая форма физиологической изоляции вполне реальна (часто у насекомых). У высших животных она приобретает формы психической изоляции. У птиц она связывается главным образом со зрительными впечатлениями от «видовых отметин», частью со слуховыми восприятиями (пение птиц). У млекопитающих основную роль играет расхождение в пахучих железах, что связано с высоким развитием органа обоняния и с его ролью в половом возбуждении. И здесь условия взаимности и согласованности в изменениях видовых отметин (окрасок, пахучих желез) с изменениями в инстинктах и поведении несовместимы с представлением об единичных мутациях или даже о простейших их комбинациях. И эти изменения мы должны рассматривать как результат эволюционного процесса, не начинающего, а завершающего видообразование. Поэтому мы оставляем здесь эти классические формы физиологической или половой изоляции без рассмотрения,— хотя возникновение половой изоляции и падает иногда на расовые различия, как это доказано, например, для двух рас (А и В) Drosophila pseudoobscura (Lancefield, Boche, по Добжанскому, 1937),— а фиксируем все наше внимание на других формах изоляции, которые, быть может, имеют несколько большее значение если не в самом возникновении расовых различий, то по меньшей мере в их относительно раннем фиксировании в качестве жизненной, в данных условиях, комбинации наследственных изменений. В этом смысле имеют значение биохимическое и генетическое расхождения, которые могут быть обозначены как гаметная и генетическая формы изоляции.
Гаметная изоляция. Межвидовая гибридизация остается обычно безрезультатной и при отсутствии каких-либо механических препятствий и даже при искусственном осеменении животных или опылении растений. У растений пыльца на чуждом рыльце не прорастает, пыльцевая трубочка не проникает в ткань столбика, не доходит до зародышевого мешка или, даже при ее успешном прорастании, генеративная клетка все же гибнет, оставляя яйцеклетку неоплодотворенной. У животных сперматозоид не проникает в яйцо или гибнет в его протоплазме. Иногда яйцо начинает дробиться, но хромосомы сперматозоида не участвуют в дроблении, выбрасываются из ядра и дегенерируют.
Во всех этих случаях мы имеем явные выражения какой-то, очевидно биохимической, несовместимости сперматозоида с тканями чуждого организма и несовместимости ядра сперматозоида с плазмой яйцеклетки. В других случаях оплодотворение совершается и дробление начинается нормально, но зародыш гибнет. У животных это происходит, по-видимому, чаще всего непосредственно после гаструляции (во время которой протекают важнейшие детерминационные процессы). В этом случае биохимическое расхождение, очевидно, не столь велико, чтобы вызвать гибель ядерных субстанций в чуждой плазме яйца, но все же настолько значительно, что их взаимодействие в процессах обмена (и в- стоящих с ними в связи детерминационных процессах) невозможно. Иногда гибридные зародыши вначале развиваются нормально и гибнут на более поздних стадиях, либо дают начало очень слабой молоди. Смертность среди таких гибридов очень велика.
Все это представляет очевидный результат биохимического расхождения организмов, в котором нарушается нормальное взаимодействие ядерных веществ и плазмы, а следовательно — нарушается и нормальное течение процессов обмена, лежащих в основе индивидуального развития и всех вообще жизненных процессов организма. Поскольку это расхождение проявляется уже в большей или меньшей несовместимости половых клеток, можно говорить о гаметной изоляции.
Совершенно ясна постепенность этого расхождения, которое проявляется в полной мере лишь в межвидовой гибридизации (да и то не всегда — в некоторых случаях возможны и межвидовые гибриды и даже увеличение мощности межвидовых гибридов — явления гетерозиса). Тем большее значение имеет для нас то обстоятельство, что начальные фазы биохимического расхождения замечаются иногда также в расовых признаках.
По Дикте (Pictet, 1936), жизнеспособность гибридов между более отдаленными географическими расами дубового шелкопряда (Lasiocampa quercus) заметно снижена. То же самое наблюдалось еще резче при скрещивании местных рас другой бабочки (Nemeo-phila plantaginis), у которых иногда гибридизация совсем не удавалась вследствие отсутствия оплодотворения. Особенно интересно, что такое снижение жизнеспособности наблюдается иногда и при скрещивании различных мутаций дрозофилы. Некоторое снижение жизнеспособности видно на комбинации abnormes — lozenge у Drosophila funebris (Тимофеев-Ресовский, 19356). Более резко это выражено в комбинации curly-wing — moire eye (Мёллер).
Очень часто расхождение двух форм выражается в том, что у гибридов недоразвиваются половые железы и половые клетки в них не развиваются или дегенерируют. Так, например, даже при скрещивании различных рас Drosophila pseudoobscura (А и В), именно — при оплодотворении самки В самцом А, получаются стерильные самцы с редуцированными семенниками. Тщательный анализ таких, а также и обратных скрещиваний (когда получаются стерильные самцы с семенниками нормальной величины), проведенный Ф. Добжанским, показал, что эти дефекты являются результатом взаимодействия наследственного материала двух разных рас (при участии не менее 8 генов «стерильности»). При скрещивании различных рас непарного шелкопряда (Lymanthria dispar) получаются, как известно (Гольдшмидт, 1935), различные интерсексы, т. е. гибриды с ненормальным половым аппаратом. И здесь, как показал Р. Гольдшмидт, нарушения в развитии половых органов объясняются взаимодействием различного наследственного материала у гибридов «слабых» и «сильных» рас этого шелкопряда. В этих последних случаях, когда недоразвивается половая система, мы имеем переходы к следующей форме — генетической изоляции, для которой характерна стерильность развивающихся гибридов. Именно в этих явлениях мы имеем начальные фазы расхождения. И в этом опять подчеркивается значение наследственных изменений ядерных структур во всех процессах внутривидового дифференцирования. Несовместимость ядра и плазмы сказывается, обычно, лишь на межвидовой или даже межродовой гибридизации, и это заставляет нас еще раз вспомнить сказанное ранее об изменчивости и, следовательно, малой устойчивости ядерных структур и относительно большей устойчивости плазмы.
Генетическая изоляция. Еще Федерлей (Federley, 1913–1916) показал на целом ряде скрещиваний различных видов бабочек (виды Pygaera и др.), что стерильность гибридов обусловлена расстройством редукционных делений при образовании половых клеток. Во время мейозиса чуждые хромосомы, все или некоторые, не спариваются и не образуют нормальных бивалентов. Эти данные были затем подтверждены и дополнены другими исследованиями над межвидовыми гибридами как у животных, так и у растений. Неконъюгирующие хромосомы (униваленты) распределяются случайно, так что половые клетки получают ненормальное и притом различное число хромосом. Важно отметить, что уже само накопление генных различий ведет к уменьшению синаптиче-ского сродства хромосом (Мёллер). Керкис (1936) нашел, что у гибридов D. melanogaster X D. simulans гомологичные хромосомы конъюгируют (в слюнных железах) совершенно даже в тех участках, которые имеют одинаковое строение. То же найдено у гибридов Drosophila virilis X D. americana (Hughes, no Muller, 1940).
Очевидно, ослабление притяжения связано с различиями в самих генах. Однако дело не исчерпывается одними лишь ненормаль-ностями в конъюгации и распределении хромосом. Клетки с нарушенным «балансом» хромосом вследствие ненормального их распределения оказываются нежизнеспособными. Что здесь дело именно в нарушении баланса, видно из того, что если у межвидовых гибридов произойдет удвоение числа хромосом, так что каждая хромосома получает свою пару, то такие «аллотетраплоиды», имеющие полные наборы хромосом, оказываются вполне плодовитыми и бойкими при внутреннем размножении. Впрочем, иногда хромосомы образуют правильные пары, но все же проявляются ненормальности в процессах редукционного или эквационного деления, и половые клетки после этого дегенерируют (иногда они и вообще недоразвиваются и не доходят даже до редукционных делений, как мы отмечали при рассмотрении гаметной изоляции). Все это относится к межвидовым гибридам. Нас интересует здесь, однако, лишь начальная дифференциация форм внутри вида и, следовательно, для нас имеет особое значение вопрос о возникновении механизма генетической изоляции и о его возможной роли в этом процессе внутривидовой дифференциации.
Ф. Добжанский (1937), который обстоятельно разработал вопрос о возникновении стерильности, различает генную и хромосомную стерильность. Нам незачем входить в особые детали этого вопроса, однако отметим, что иногда стерильность определяется изменением одного лишь гена, который обнаруживает типичное менде-лирование при скрещивании. Несколько таких генов описано Бидлем у кукурузы (Beadl, 1930–1933). Описаны такие гены и для видов дрозофилы (Sturtewant, 1929 а, б; Gowen, 1931). Здесь отдельная мутация выражается в нарушениях конъюгации хромосом, в их неправильном расхождении или в других ненормально-стях при образовании половых продуктов. В высшей степени интересны данные Мёллера и Раффеля, показавшие, что при скрещивании двух вполне плодовитых мутаций дрозофилы curly-wing и moire eye получаются слабые и бесплодные гибриды. То же самое наблюдается при скрещивании scute-18 и scute-4 (Muller, 1940). Здесь получаются, следовательно, стерильные комбинации о т-дельных мутаций. Чаще всего, по-видимому, стерильность определяется не отдельной мутацией, а несовместимостью целых генотипов, которые дают негармоничную комбинацию, ведущую, по меньшей мере, к бесплодию. В других случаях стерильность определяется несовместимостью хромосом вследствие наличия крупных структурных различий — именно транслокаций или более сложных инверсий. Расы А ж В D. pseudoobscura различаются четырьмя инверсиями. При скрещивании получаются стерильные самцы. Кожевников (1936) получил расу дрозофилы {D. melanogaster), отличающуюся двумя транслокациями (D. artificialis). При чистом разведении она обладает лишь сниженной плодовитостью (жизнеспособна lU часть всех гамет). Скрещивание с исходной нормальной дрозофилой остается безрезультатным, так как оно не может привести к образованию зиготы со «сбалансированной» системой хромосом.
Генетики считают, что в основе стерильности гибридов лежит либо изменение некоторых генов, т. е. отдельные мутации «стерильности», которые обнаруживают свое влияние в чуждом генотипе, либо негармоническая комбинация генов или хромосом (т. е. нарушение их «баланса»).
Хотя физиологическая изоляция является, очевидно, дальнейшим и притом обычно побочным результатом эволюции, интересно отметить, что все же она и сама по себе может быть результатом непосредственного действия естественного отбора. Два близких вида дрозофилы D. pseudoobscura и D. miranda могут быть скрещены, но дают стерильных гибридов. Между ними имеется известная физиологическая изоляция — в смешанных популяциях (в эксперименте) преобладают, особенно первое время, внутривидовые спаривания. Эта изоляция различно выражена для различных рас (раса В. у D. pseudoobscura) и линий (D. miranda) l, которые обитают в одном и том же ареале, и значительно ниже для тех рас и линий, которые происходят из отдаленных местностей (Добжанский, 1937). Очевидно, шел естественный отбор на физиологическую изоляцию только тех популяций различных видов, которые, живя в одной местности, фактически могли встречаться. Этот отбор основан на элиминации потомства особей, которые скрещивались с особями другого вида (так как это скрещивание вело к образованию бесплодных гибридов).
Накопление генных мутаций и хромосомных перестроек ведет к уменьшению сродства, выражающемуся в уменьшении притяжения между гомологичными хромосомами. Оно ведет к нарушению редукционных делений и к другим проявлениям несовместимости наследственного аппарата расходящихся форм. Нарушение «баланса» генов и хромосом является таким же проявлением их несовместимости в процессах обмена веществ и в индивидуальном развитии организма. Оно ведет к «летальности» или «стерильности» несовместимых комбинаций. Несовместимость ядра и плазмы является дальнейшим и еще более глубоким проявлением биохимического расхождения различных форм в процессе эволюции. Все эти процессы способствуют все более полному их обособлению.
Колебания численности популяций (периодическое ограничение панмиксии). Мы уже говорили вначале о роли чередования различных форм размножения, связанных с периодическим прекращением панмиксии. У высших животных такое чередование поколений не играет никакой роли (за немногими исключениями), но зато у многих животных наблюдается временное ограничение панмиксии, связанное с более или менее резким сокращением численности. Мы знаем, что панмиксия осуществляется более свободно лишь в больших популяциях подвижных животных. В малых популяциях преобладает родственное скрещивание, ведущее к гомозиготизации и закреплению различных случайных комбинаций наследственных изменений. В первом случае (в больших популяциях) свободная панмиксия позволяет осуществить много новых комбинаций, но не дает возможности их закрепления. Во втором случае (в малых популяциях) родственное скрещивание ограничивает возможность комбинирования, связано даже с утерей многих уже осуществленных комбинаций, но, с другой стороны, может привести к фиксированию и размножению некоторых благоприятных сочетаний. В частичной пространственной изоляции отдельных популяций широко распространенного вида происходит известное совмещение преимуществ большой популяции, создающей новые комбинации благодаря возможности миграций, и выгод, связанных с дифференциацией на малые популяции, что способствует закреплению удачных комбинаций.
При периодических колебаниях численности также происходит подобное совмещение обоих условий видообразования путем чередования процессов комбинирования и процессов фиксирования и размножения.
Колебания численности могут носить несезонный характер, и именно эти колебания имеют большое народнохозяйственное значение, так как их нелегко предусмотреть. Факторы, способствующие массовому размножению, и факторы, определяющие наступление следующего за ним периода депрессии, могут быть весьма различными. Это могут быть случайные изменения в физических факторах (температура, влажность), прямо или косвенно благоприятствующих размножению или, наоборот, ограничивающих это размножение, изменения в условиях питания (неурожаи кормов) или изменения в биологических соотношениях (истребление хищниками, истощение паразитами, эпизоотии). Иногда выясняется и вполне закономерная зависимость в колебаниях численности различных организмов, объединенных пищевыми связями. Известны сопряженные изменения численности зайца и рыси в Канаде (Hewitt, 1921) и многие другие связи, прослеженные на протяжении многих лет главным образом по масштабам заготовок шкурок пушных зверей, а также по биологическому учету численности (см. сводки С. А. Северцова, 1941; А. Н. Формозова 1934; Elton, 1924, 1927). Несезонные колебания численности насекомых, а отчасти грызунов и даже крупных млекопитающих могут определяться и метеорологическими условиями — случайно особо благоприятная весна может привести к интенсивному размножению насекомого, дождливое лето может значительно снизить его численность, а глубокий снежный покров и, в особенности, его обледенение могут привести к длительной голодовке и массовой гибели даже крупных травоядных млекопитающих. Это все — наиболее простые случаи. Чаще всего, однако, у мелких животных (насекомых), при сложности связей и малой специализации питания, массовое размножение или почти полное исчезновение зависит от сложного сочетания многих благоприятных или неблагоприятных факторов.
Для процессов эволюции имеют, пожалуй, еще большее значение почти всеобщие для умеренных и холодных стран явления сезонных колебаний численности. Во время зимовок ежегодно гибнет большое число птиц и млекопитающих и огромное количество насекомых. Отношение численности популяций перед зимовкой и после нее достигает 1000:1 или более. Разница в величинах осенней и весенней популяций бывает очень значительной и у птиц, и у мелких млекопитающих.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 554; Нарушение авторских прав?; Мы поможем в написании вашей работы!