КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Изменение выражения мутаций в процессе эволюции
|
|
|
|
Не только мута|бильность (мутационное «давление»), но и отборное значение, коэффициент селекции) признаков не остаются постоянными. И это объясняется не только постоянными изменениями факторов среды, но и изменениями самого организма. Даже отдельная мутация не имеет постоянного выражения (рис. 19). Поэтому и концентрация мутаций не устанавливается на каком-либо уровне равновесия. Этот уровень непрерывно меняется, по меньшей мере — для всех небезразличных признаков.
Так как отдельные мутации вносят обыкновенно неблагоприятные изменения организации (с ее функциями), то нелегко было бы понять, каким образом возможна прогрессивная эволюция, если бы мы не владели громадным арсеналом фактов, показывающих, что неблагоприятные выражения мутаций в процессе эволюции погашаются и преобразуются. Это достигается в результате комбинирования различных мутаций при скрещиваниях и естественном отборе наиболее жизненных сочетаний.
Генетиками уже давно было установлено, что фенотипическое выражение мутаций может резко изменяться при их комбинировании. Классическим примером такого комбинирования являются различные формы гребня у петухов (рис. 20). В данном случае, как и во многих других, наблюдается именно коренное преобразование выражения единичных мутаций. Комбинация дает качественно новое формообразование. В других случаях мутация получает известное выражение вообще лишь в комбинации с определенной другой мутацией — в отдельности мутация не проявляется. Такие факторы генетики называют дополнительными («комплементарными»). Так, например, основной «ген» окраски у домашней мыши определяет развитие серой или черной окраски лишь в присутствии дополнительных факторов. Известны также дополнительные факторы, определяющие развитие окраски цветов душистого горошка. Наконец, в третьих случаях, наоборот, мутация полностью теряет свое выражение при комбинировании с определенной другой мутацией, которая оказывает «эпистатическое» влияние на первую. Классическим примером такого эпистаза может служить доминантная белая окраска итальянских кур (леггорнов), которая подавляет развитие иных окрасок.
Эти примеры относятся к крайне резкому проявлению эффекта комбинирования двух простых мутаций. Выражение одной мутации проявляется лишь при наличии другой (комплементарной) или, наоборот, полностью подавляется (эпистатическим фактором). Все ото указывает на наличие известного взаимодействия, конечно — не непосредственно между «генами», как это иногда упрощенно описывают, а между теми биохимическими и морфогенетическими процессами, которые оказались измененными в результате данных мутаций.

Рис. 20. Гребни домашних кур. Доминирование уклоняющихся форм гребня — гороховидного (А) и розовид-ного (В)—над простым (С). Комбинирование гороховидного и розовидного гребней дает качественно новую ореховидную (D) форму гребня. По Р. Пеннет и П. Бейли, 1921
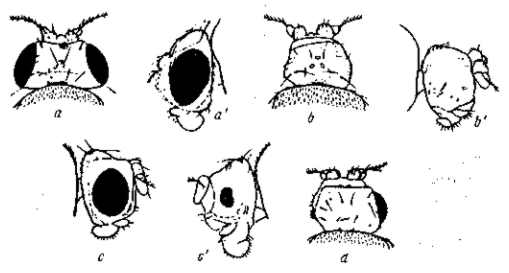
Рис. 21. Изменчивость выражения мутации безглазия (eyeless) у Drosophila melanogaster. а, а' — нормальные глаза; b, b' — совершенно безглазая форма. В чистой культуре в течение ряда поколений выражение этой мутации постепенно теряется (с, с') вплоть до восстановления почти нормальных размеров глаза ) и нормальной жизнеспособности мутантных мух. По Т. Моргану, 1916
Мы сослались на известные всем генетикам типичные примеры такого взаимодействия. В некоторых случаях модифицирующее влияние одних мутаций на другие выражается исключительно ярко. В менее резкой форме подобное взаимодействие не составляет исключения, нуждающегося в определенных терминах, а является обычным правилом. Меняется выражение и в компаундах разных аллеломорфных мутаций. Иногда, правда, бывает, что каждая мутация сохраняет свои особенности и в компаунде (например, aristopedia X spineless). Чаще, однако, эти выражения меняются. Интересно, что нередко выражения обоих аллеломорфов взаимно погашаются. Так, vortex (вихреобразные щетинки) и oblique (косо срезанные крылья) дают в компаунде почти нормальный фенотип. То же самое касается split bristles X Notch, а также achaete X scute-1, теряющих в компаунде свои выражения (Мёллер).
«Доминантная» мутация четковидных крыльев (Beaded) обладает в гетерозиготном состоянии крайне изменчивым выражением, меняющимся как при комбинировании, так и при изменении внешних факторов (рис, 19). Фактор black (черное тело) даже в гетерозиготном состоянии подавляет до известной степени выражение Beaded, а мутация vestigial (рудиментарные крылья) в гетерозиготном же состоянии усиливает его эффект. Мутация sooty оказывает также усиливающее влияние на выражение мутации четко-видных крыльев (Мёллер). При выращивании личинок этой мутации в подсохшей среде четковидность оказывается менее выраженной и часто вовсе не проявляется, так что значительная часть мух оказывается фенотипически нормальной. Так же влияют и различные другие внешние факторы (Декстер, 1914). Большая зависимость от генотипа установлена и для выражения мутации Beaded в гетерозиготном состоянии (Шифрин, 1941).
Рецессивные мутации рудиментарных крыльев (vestigial), а также безглазия (eyeless) значительно изменяются в зависимости от температуры (рис. 21, 22) и от качества корма. Мутация безглазия также весьма чувствительна и по отношению к модифицирующим влияниям со стороны других мутаций. Однако иногда изменчиво даже выражение «доминантной» мутации в гомозиготном состоянии. Так, мутация уродливого брюшка (Abnormal abdomen) дрозофилы в своем выражении весьма зависима от влажности корма, а мутация полосковидных глаз (Ваг) весьма чувствительна к изменениям температуры (рис. 6).
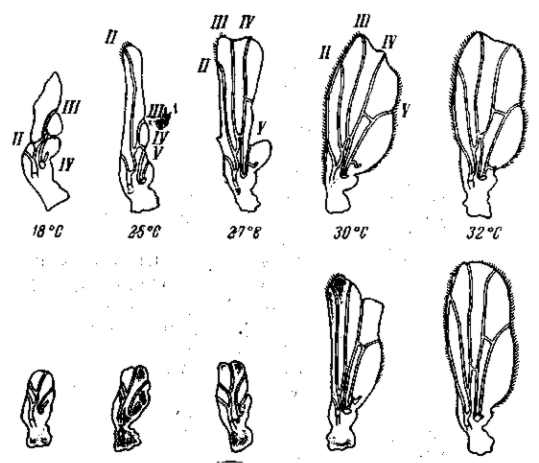
Рис. 22. Зависимость формы и величины крыльев мутации vestigial (Drosophila теlanogaster) от температуры Верхний ряд — самцы, нижний — самки. При 32° в культуре этой мутации 18% самцов и 15 % самок имеют совершенно нормальные крылья (проявление становится неполным). По М. Г. и M. Л. Харнли, 1935
Как мы раньше уже отмечали, каждая мутация выражает либо изменение времени, скорости образования и концентрации морфо-генных веществ, либо изменение реактивности тканей (сдвиг пороговых уровней нормальных реакций). В обоих случаях это означает взаимное смещение формативного раздражителя (активатора) и реагирующей системы (реактора). В огромном большинстве случаев смещение приводит также к уменьшению «морфогенного запаса» и приближению измененной интенсивности активатора (индуктора), к одному из порогов нормальной реактивности реактора. Такое приближение естественно ведет к уменьшению устойчивости формообразовательного процесса. Близость порогового уровня означает незащищенность морфогенетической реакции достаточным «запасом». Самое незначительное дальнейшее уклонение в том же направлении,— либо под влиянием небольших наследственных изменений (малых мутаций), либо под влиянием изменения внешних факторов,— может привести к превышению порога нормальной реактивности и к более или менее значительному нарушению формообразования. Поэтому те мутации, которые стоят как раз на грани подобных нарушений, оказываются особенно неустойчивыми в своем проявлении (т. е. обладают неполным проявлением) или в своем выражении.
Полностью рецессивные мутации в гетерозиготном состоянии не проявляются, т. е. в однократной «дозе» при наличии нормального аллеломорфа вызывают сдвиг, не достигающий порогового уровня нормальной реактивности тканей. В гомозиготе они проявляются, и это указывает на то, что двукратная «доза» изменения «гена» выводит взаимодействующую морфогенную систему за пределы порога нормальной реактивности. Если она оказывается как раз на грани этого порога, то такая мутация будет в своем проявлении и выражении весьма неустойчивой. Она окажется в широких пределах зависимой как от наличного генотипа, так и от внешних влияний. Возможна мутация, которая не проявляется и в гомозиготе, но все же вызывает заметный сдвиг нормальной реактивности, который при суммировании с другой мутацией приведет к видимому изменению фенотипа или к изменению его физиологического состояния. Это — «малая» мутация, выступающая лишь в роли «модификатора».
С другой стороны, возможны более эффективные мутации, которые не только в двукратной дозе, но и в гетерозиготе, при наличии нормального аллеломорфа, достигают или даже превышают допустимые пределы сдвига компонентов взаимодействующей системы. Если сдвиг достиг порогового уровня, то получится картина доминирования с неполным проявлением (в зависимости от генотипа и внешних факторов). Если этот пороговый уровень полностью превзойден, то возможно устойчивое проявление данной мутации при изменчивости ее выражения.
Ни об одной мутации не известно, чтобы она достигла того уровня устойчивости, каким обладает нормальный организм. Это означает, что если порог «нормальной» реактивности и превзойден полностью (как в доминантных мутациях), то все же не настолько, чтобы создать «запас», «защищающий» формообразование мутации. Такие «запасы» создаются исключительно в процессе эволюции (т. е. при направляющем влиянии отбора), а не в порядке единичных мутаций. Они «защищают» развитие уже установившейся «нормы» («фактор безопасности» Холдена).
Мутации никогда, по-видимому, не достигают уровня полной доминантности (пока они остаются в роли мутаций). То, что у мутаций дрозофилы называется доминированием, никогда не достигает высоты доминирования нормального фенотипа. Доминирование нормы выражается обычно в почти полной или полной неотличимости гетерозиготы от гомозиготы (по фенотипу). Между тем при доминировании мутации гетерозигота всегда легко отличима от гомозиготной мутации. Последняя обладает гораздо более ярким выражением характерных для нее признаков. Гетерозигота доминантной мутации занимает всегда промежуточное положение между нормой и гомозиготной мутацией. Таковы все известные доминантные мутации дрозофилы. Одним словом, доминантные мутации на самом деле всегда бывают полудоминантными.
Полудоминанты отличаются неустойчивостью именно потому, что они не достигли уровня настоящих доминантов, т. е. той активности, которой отличаются «нормальные» аллеломорфы. Вернее — измененные в мутациях формообразовательные процессы никогда не обладают той «защищенностью», которая характерна для процессов индивидуального развития нормального «дикого» типа. Это связано с кратковременностью их исторического существования и отсутствием положительного селекционного значения данной мутации.
Мы видим, что проявление и выражение мутаций, как в гетеро-зиготе, так и в гомозиготе, суть величины переменные. Они могут изменяться как в сторону усиления, так и в сторону ослабления. В этом смысле имеют значение и внешние факторы среды, в которой развивается организм (мутация), и внутренние факторы, т. е. генотип данной особи.
В процессе эволюции особое значение имеет комбинирование. При скрещиваниях создаются все новые комбинации, а вместе с тем меняются выражения мутаций, меняются фенотипы и их реакции (рис. 15, 16). Так как большинство мутаций так или иначе неблагоприятно, то наибольшее значение имеет частичное или полное погашение их выражения.
А. ОБЕЗВРЕЖИВАНИЕ НЕБЛАГОПРИЯТНЫХ МУТАЦИЙ (ЭВОЛЮЦИЯ РЕЦЕССИВНОСТИ)
Неоднократно делались попытки повлиять путем искусственного отбора на мутации с изменчивым выражением. Эти попытки давали положительные результаты. Путем искусственного отбора удавалось значительно поднять выражение мутации до известного максимума и удавалось снизить его, иногда — до уровня дикой нормы, т. е. до полной утраты проявления. Эти изменения оказывались наследственными и зависели, следовательно, от отбора небольших наследственных изменений (малых мутаций), меняющих выражение культивировавшихся мутаций. Такие малые мутации были названы «модификаторами». В этом «модифицировании», конечно, нет ничего специфического,— мы ведь видели, что взаимодействие «генов» (точнее — измененных биохимических и морфо-генетических процессов) вообще связано с изменением выражения отдельных мутаций. В термине «модификатор» подчеркивается, однако, модифицирующее влияние только потому, что во многих малых мутациях нам не удается пока уловить их влияния на нормальный организм (так как «дикая» норма отличается гораздо большей устойчивостью, т. е. «защищенностью» онтогенеза). Поэтому их наличие пока устанавливается только по влиянию на выражение мутаций и то главным образом лишь по комплексному влиянию целого ряда малых мутаций.
Анализ фенотипов любой природной популяции показывает, обычно, поразительное единообразие. В частности, в популяциях различных видов дрозофилы, в том числе и классической Drosoрhila melanogaster, решительно господствует нормальный «дикий» тип. И все же генетический анализ показывает огромную их насыщенность разнообразнейшими мутациями, в том числе и полудоминантными. Все эти мутации находятся, однако, в скрытом виде, большей частью в качестве рецесссивов (в гетерозиготном состоянии; С. Четвериков, Н. Дубинин и др.). Однако встречаются не так уж редко и полудоминанты — выражение их частью подавлено эпистатическими влияниями генотипа или же они обладают низким процентом проявления (Р. Берг, С. Гершензон). Встречаются и довольно ясно выраженные мутации (по Р. Берг, особенно много полудоминантных аутосомных мутаций) и притом не безвредные. Имеется в природных популяциях и немало летальных (Н. Дубинин, Ф. Добжанский), а также стерильных (Р. Берг, 938) мутаций. Однако в особенности велико насыщение малыми мутациями, выступающими в роли модификаторов (Н. Дубинин и др.), и физиологическими мутациями, отражающимися лишь на относительной жизнеспособности (Г. Муретов, 1941). Это показывает, что в природных условиях происходят те же процессы насыщения популяций мутациями и их обезвреживания путем отбора наиболее благоприятных комбинаций, т. е. прежде всего отбора наиболее «жизнеспособных» особей, какие установлены для лабораторных культур.
И в природных условиях неблагоприятные мутации теряют свое выражение. В основном это касается гетерозиготы, так как гомозиготные мутации могут встретиться в природе лишь в виде исключения. Результатом утраты своего выражения в гетерозиготе является рецессивность мутации.
То, что нами наблюдается в лаборатории и в природных условиях, является лишь ничтожным отрезком истории организмов. Те мутации, которые мы культивируем как вновь возникшие наследственные изменения и на которых мы наблюдали изменение их выражения в течение исключительно короткого времени, возникали многократно и ранее в истории данного вида организмов (например, дрозофилы). Организм уже до известной степени приспособился к ним, т. е. выработал различные компенсации, подавляющие их неблагоприятные выражения. В частности, под этим углом зрения можно оценивать обычные явления доминирования нормы и рецессивности мутаций.
В действительности, нормальный фенотип более или менее полно доминирует над всеми мутациями. Мы уже отмечали, что даже немногие известные «доминантные» мутации дрозофилы в действительности являются полудоминантными. Их выражение в гетерозиготе всегда ослаблено по сравнению с гомозиготой. Частично это может объясняться тем, что большинство мутаций связано с ослаблением интенсивности нормальных процессов развития (замедлением продукции и снижением концентрации морфогенных раздражителей или снижением реактивности тканей, т. е. поднятием нижнего порога и снижением верхнего порога чувствительности) и ведет к известному недоразвитию или упрощению морфогенеза. Генетики говорят об инактивировании «генов» и «гшгоморфных» или «аморфных» генах (Мёллер, 1932).
Однако уже факт существования обратных мутаций показывает возможность противоположных изменений в сторону интенсификации морфогенетических процессов («гиперморфные» гены Мёллера — ускорение образования и повышение концентрации формообразовательных веществ, а также повышение чувствительности тканей, т. е. раздвигание обоих пороговых уровней их реактивности). Возможны и такие изменения, которые противодействуют течению нормальных реакций или вносят в них нечто совершенно новое (антиморфы и неоморфы Мёллера). В этих случаях наблюдается доминирование мутации. Однако, как уже сказано, это доминирование в действительности никогда не бывает полным. Можно предполагать, что выражение этих мутаций также уже частично ослаблено. Вместе с тем нельзя не отметить, что таких мутаций известно очень немного. Огромное большинство мутаций дрозофилы — рецессивно. Очевидно, это объясняется так же, как и в прошлой истории вида: при повторном возникновении этих, не слишком вредных, мутаций шел естественный отбор комбинаций, нейтрализующих их выражение. Такие комбинации (генотипы) отличались большей жизнеспособностью ираспространялись в популяции в качестве все более устойчивой «нормы».
Одновременно с развитием генетического механизма, обезвреживающего наиболее частые мутации, происходило упрочение процессов нормального формообразования даже гетерозиготного организма, т. е. усиление доминирования нормы, и вместе с тем ослабление выражения неблагоприятных мутаций (в той же гетерозиготе) вплоть до их полного погашения, т. е. приближение к полной рецессивности мутаций.
Теория эволюции доминантности Фишера построена на значении отбора малых мутаций с модифицирующим влиянием. Предполагается, что каждая действительно новая мутация обладает всегда некоторым выражением и в гетерозиготном состоянии, т. е. все вновь возникающие мутации полудоминантны. Если такая мутация непосредственно не элиминируется, т. е. по своему выражению не слишком неблагоприятна (при данных условиях существования), и оставляет какое-то потомство, то она комбинируется в этом потомстве с несколько различными генотипами. Выражение ее оказывается изменчивым, и в процессе естественного отбора переживают всегда наиболее приспособленные, наиболее жизнеспособные комбинации. Если такой процесс многократно возобновляется и поддерживается мутированием и распространением мутации, то постоянный отбор в пользу нормального фенотипа приводит к накоплению в данной популяции «модификаторов», нейтрализующих вредные выражения распространенных мутаций. Тем самым эти мутации становятся рецессивными в данной популяции, насыщенной соответствующими модификаторами, а норма — по отношению к ним — доминантной.
Если среди новых полудоминантных мутаций встречается относительно (в данных условиях) благоприятное изменение, то его выражение будет через тот же отбор «модификаторов» (т. е. попросту через отбор более приспособленных особей) постепенно усиливаться, вплоть до максимально возможного выражения в гетерозиготе. Это означает приобретение полной доминантности.
Благоприятная доминанта быстро распространится при участии положительного отбора, перейдет в гомозиготное состояние и войдет в состав новой нормы, вытесняя прежнюю норму; прежний нормальный аллеломорф потеряет свою доминантность и в качестве рецессива перейдет в скрытое состояние.
Таким образом, большинство не слишком вредных мутаций становится рецессивным. Однако такие мутации, которые заметно ослабляют жизнеспособность и приспособленность организма к данным условиям внешней среды, не имеют шансов переживать и оставлять потомство. Ясно, что такие постоянно элиминируемые мутации сохраняют всегда свои исходные свойства «новых» мутаций, т. е. остаются полудоминантными, так как никакой отбор среди нежизнеспособных особей невозможен. В результате целый ряд мутаций дрозофилы оказывается полудоминантным. В особенности это касается ряда более значительных физиологических мутаций с летальным или стерильным эффектом.
Другие авторы, присоединяясь к основным положениям Фишера об эволюции доминантности, представляют себе этот процесс несколько иначе. Райт, Холден и Мёллер считают, что доминантность выражает активность факторов нормального развития. Обычно при этом говорят об активности нормальных «генов» и об инактивации мутантных (в большем или меньшем согласии с представлениями Гольдшмидта о количественном характере изменений генов и об их энзиматозной природе). Нам незачем связывать вполне реальные данные об изменении активности или концентрации формативных веществ и об изменении времени и скоростей морфогене-тических реакций с гипотетическими представлениями о природе гена, о характере его изменений и о способе его участия в индивидуальном развитии организма. Я пытался показать (И. Шмальгаузен, 1938а, 1942), что почти все мутации, вызывая сдвиги в мор-фогенетических системах (между «активатором» и «реактором») организма, вносят известное нарушение развития, которое при превышении порогового уровня известной реакции вызывает выпадение этой реакции. Такие выпадения некоторых реакций должны выражаться в большей или меньшей степени недоразвития. Обычно мутация как раз и связана с известным недоразвитием, т. е. с остановкой развития или заменой сложного онтогенетического процесса более простым. Естественно, что такие мутации имеют характер «гипоморфньгх» мутаций и в комбинации с нормальным аллеломорфом оказываются до известной степени рецессивными. Однако большая устойчивость нормы и более или менее полная рецессивность, очевидно, не объясняются так просто.
По мнению Мёллера, понижение (или повышение) степени выражения известной мутации зависит не от подбора генов — модификаторов этой мутации, а от отбора на большую (или меньшую) активность того же основного гена, который определяет развитие нормы. При наличии ряда аллеломорфных изменений, градуально друг от друга отличающихся, такой отбор вполне возможен. Следовательно, вредные мутации будут уничтожаться через отбор гетерозиготных особей с наименьшим их выражением, т. е. наибольшей активностью «нормального» аллеломорфа. Наоборот, благоприятные мутации будут повышать свою активность, т. е. увеличивать степень доминирования. Повышение активности до известного уровня приведет к тому, что уже один ген (1А) даст полную реакцию в виде реализации характерных признаков в их максимальном (оптимальном) выражении, так что его удвоение (2А в гомозиготе) не даст уже никакого дополнительного эффекта. В таком случае будет достигнуто полное доминирование. Те мутации, которые имеют положительное значение для жизни вида, будут всегда подбираться по этому пути повышения активности вплоть до развития полной доминантности. Полезная мутация, став доминантной, распространится с большой скоростью по всей популяции и войдет в состав ее нормы.
Мутации, до известной степени вредные, будут переходить в менее активное состояние и в обезвреженном виде могут сохраняться в популяции в виде скрытого резерва ее изменчивости (в гетерозиготах).
По представлениям Райта, степень доминирования есть также одно из выражений активности гена, и рецессивность означает его переход в менее активное состояние. Холден не отрицает значения модификаторов, действующих на мутацию. Холден добавляет к этому, однако, еще и следующие соображения. Если нормальный ген вызывает уже в гетерозиготе полный эффект, то всякое увеличение его активности останется незамеченным. Мутация с повышенной активностью («гиперморфы» Мёллера), которые могли бы быть полностью доминантными, не получают видимого выражения. Только минус-мутация может получить свое выражение, если активность гена в гетерозиготе падает ниже нормы, т. е. если она рецессивна. Такие мутации обычно менее жизнеспособны и частью элиминируются, частью видоизменяются в процессе эволюции. С другой стороны, плюс-мутации, связанные с переходом «гена» в более активное состояние, хотя и не дают видимого эффекта, но ведут к увеличению устойчивости нормы. Они свободно накапливаются в процессе отбора. Поэтому отбор всегда будет действовать в сторону повышения активности (и доминирования) всех жизненно важных генов.
Исследования результатов межвидовой гибридизации некоторых видов хлопчатника показали, что доминантность имеющихся у них параллельных мутаций зависит от различных (у разных видов) комплексов модификаторов. Они приобретены в истории данных видов независимо друг от друга и имеют поэтому разный состав. При межвидовом скрещивании доминирование оказывается лишь слабо выраженным вследствие очевидной дезинтеграции обеих систем модификаторов, определяющих это доминирование (Харланд, 1936, 1937). В особенности ясно это проявляется при внесении мутации в чуждый ей вид путем повторного возвратного скрещивания с этим видом. Это является прекрасным доказательством правильности теории эволюции доминантности в том ее виде, как она была разработана Фишером (1931). Доказательствами эволюции рецессивности являются и следующие данные того же Харланда. Фактор crinkled — dwarf оказывается простым рецессивом в одном сорте хлопка, однако в скрещивании с другими сортами хлопка, т. е. в других комбинациях генов (включая «модификаторы»), он обнаруживает во втором поколении, полученном путем самоопыления гибридов, все степени доминирования. По Фишеру, это объясняется расщеплением в отношении многочисленных модификаторов. Однако и в этих случаях допустимо не только предположение о дезинтеграции системы модификаторов, обусловливающих рецессивность данной мутации. Можно допустить и другие толкования, в особенности потому, что эта же мутация оказывается вполне рецессивной и в хлопчатнике «упланд» (Gossypium hirsutum), в котором она никогда не встречается и который, по Фишеру, не может обладать системой модификаторов, обусловливающих ее рецессивность.
В данном случае эти детали не имеют большого значения, и нам представляется более правильным считать, что как в процессе эволюции вообще одинаковые результаты достигаются самыми различными средствами, так и в данном случае весьма вероятно, что эволюция рецессивности (и доминантности) идет разными путями. Повышение устойчивости нормы как путем отбора модификаторов, так и через отбор более активных аллеломорфов, при которых достигается наибольшая надежность формообразования не только в гетерозиготе, но и в гомозиготе, представляет одну сторону общего процесса стабилизации нормы. Уменьшение выражения всех вредных мутаций в результате отбора менее активных аллеломорфов, а также отбора модификаторов и вообще различных наследственных сочетаний представляет другую сторону того же процесса стабилизации нормы, который осуществляется всегда и повсюду через стабилизирующую форму естественного отбора на основе селекционного преимущества нормы перед всеми от нее уклонениями.
Б. ИЗМЕНЕНИЯ ЧАСТИЧНО ВРЕДНЫХ МУТАЦИЙ (ВОЗНИКНОВЕНИЕ КОРРЕЛЯЦИЙ)
Так как мутации имеют обычно множественное выражение, а благоприятные выражения встречаются исключительно редко, то большое значение в эволюции имеет возможность нейтрализации одних лишь вредных выражений, а также возможность качественного преобразования других выражений и усиления положительного эффекта тех же мутаций. Такие сложные преобразования предполагают существование весьма дифференцированного действия модификаторов и приводят к установлению сложных систем связей (генетических корреляций), входящих тогда как необходимый элемент наследственной структуры организма.
Возможность дифференцированного изменения различных выражений одних и тех же частично вредных мутаций доказывается целым рядом фактов. Напомним прежде всего, что в основе мутаций лежат, очевидно, некоторые биохимические изменения, оказывающие влияние на процессы внутриклеточного обмена веществ. Только через эти изменения в обмене веществ развивающегося организма они вызывают нарушения в ходе морфогенетических процессов и приводят к видимому изменению нормального строения организма. Поэтому нужно думать, что любая морфологическая мутация имеет обязательно и физиологическое выражение. Даже малые мутации, не проявляющиеся в заметных изменениях структуры (хотя бы и в силу наличия совершенной регуляции), могут все же иметь немалое физиологическое значение. На самом деле, мы знаем, что все «вредные» мутации дрофозилы обладают ослабленной жизнеспособностью, пониженной плодовитостью, измененным темпом роста или развития, измененной реактивностью или другими физиологическими выражениями. С другой стороны, действительно известны как малые, так и большие физиологические мутации, не обладающие видимым морфологическим выражением.
Целый ряд видимых мутаций дрозофилы связан с резким снижением жизнеспособности — они «летальны» или полулегальны. Не может быть случайностью, что такие мутации оказываются в своем морфологическом выражении обычно полудоминантами, а в физиологическом выражении — рецессивами. Таковы «доминантные» мутации Notch, Dichaete, Star, Beaded, Truncate. Все они в гетерозиготе вполне жизнеспособны, а в гомозиготе оказываются полностью летальными. То же самое найдено и у амбарной огневки (Ephestia kuhniella). Мутация, сдвигающая поперечные полосы рисунка крыльев (Sy), вполне выражена, т. е. полудоминантна в гетерозиготе, и вполне жизнеспособна. Однако в гомозиготе она детальна. Известная мутация желтой окраски у домашней мыши доминантна, но обладает рецессивным летальным действием, обнаруживающимся в гомозиготе. Коротконогость кур (криперов) доминантна, но в гомозиготе она детальна. То же самое касается известных пород домашнего скота Niatta и Dexter. Последние обладают свойствами, ценимыми человеком, и потому здесь можно говорить лишь о влиянии искусственного отбора. Что же касается дрозофилы и огневки, то различная активность (доминантность) разных выражений одних и тех же мутаций является, очевидно, результатом естественного отбора.
По-видимому, морфологические выражения этих мутаций были не столь вредны, как ослабление общей жизнеспособности в гетерозиготном состоянии. Кроме того, постоянное наличие большого запаса малых «физиологических» мутаций способствовало большой эффективности отбора на «жизнеспособность» гетерози-готы, т. е. на устранение проявления признаков «летальности». Поэтому физиологическое выражение этих мутаций снижено до полной рецессивности, в то время как менее вредное морфологическое выражение осталось еще полудоминантным. Во всяком случае, это показывает возможность изменения соотношений разных выражений одной и той же мутации в гетерозиготе и в гомозиготе.
Несомненно, возможны были бы и диаметрально противоположные соотношения — погашение морфологического выражения при сохранении физиологического. Это доказывают результаты некоторых опытов комбинирования мутаций у той же дрозофилы.
Мутация lozenge обладает грубыми фасетками неправильной формы, а мутация spectacled характеризуется блестящей поверхностью глаза без фасеток. Это — аллеломорфы, которые, каждая в отдельности, нарушают строение глаза, но, кроме того, связаны с бесплодием самок. В компаунде обеих мутаций глаза получают нормальное строение, однако самки остаются бесплодными. Аллело-морфные мутации split bristles и facet также нарушают строение омматидий. В компаунде восстанавливается нормальное строение глаз, но развиваются добавочные щетинки (как в первой мутации). Черночешуйчатая мутация амбарной огневки (Ephestia kiihniella) рецессивна, но снижение жизнеспособности (на 22%) у нее доминантно (случай, обратный обычному; возможно, что это особая форма защиты организма от вредной мутации; тогда она могла развиться лишь в процессе межгрупповой конкуренции разных популяций). Эти факты показывают возможность изменения отдельных выражений мутаций в различных направлениях через процесс их комбинирования.
Добжанский (1927) указывает, что аллеломорфы белых глаз (white), а также мутации окраски тела sooty и ebony у дрозофилы рецессивны, поскольку более выраженная неблагоприятная окраска у гетерозиготы, очевидно, элиминировалась и в процессе отбора произошло восстановление нормальной, дикой окраски. Однако плейотропное выражение тех же мутаций — измененная форма сперматеки, очевидно (как нейтральный признак внутренней организации), не была предметом отбора. Она оказывается у них полудоминантной и дает в гетерозиготе промежуточные формы.
Хохлатость кур связана с гидроцефалией и недоразвитием крыши черепа. Однако эти последние выражения очень изменчивы и могут быть подавлены, как это видно из факта существования пород хохлатых кур, имеющих нормальные черепа.
Мохноногость кур связана с брахидактилией четвертого пальца. Однако и это выражение очень изменчиво, что указывает на существование многих модификаторов. В самом деле, брахидактилия может быть полностью подавлена. У некоторых пород кур (фаве-ролей) мохноногость не сопровождается брахидактилией.
В случае положительного отбора по одному выражению и элиминации особей с более значительными иными выражениями возможна, таким образом, стабилизация положительного и устранение отрицательных выражений. Это происходит через отбор мутаций с модифицирующими влияниями. Возможно установление новой нормы, обладающей известными преимуществами перед прежней. В этом случае новый благоприятный признак связан с целой системой модификаторов, совместно участвующих в его развитии. Они становятся необходимыми элементами аппарата индивидуального развития. Они действуют притом на одни биохимические реакции и на одни части морфогенетической системы как активаторы, а на другие части как депрессоры или трансформаторы реакций. Это означает установление сложных связей между различными процессами. Если уже самый факт наличия плейотропного (множественного) выражения мутации показывает наличие известных коррелятивных зависимостей между физиологическими и морфогенетическими процессами, то преобразование различных выражений этой мутации путем комбинирования со многими модификаторами означает укрепление и преобразование данной системы корреляций, а также создание новой системы геномных связей. Развитие отдельных признаков оказывается зависящим не только от основного мутант-ного «гена», но и от многих других наследственных факторов. Это выражается в наличии сложных связей между биохимическими процессами, определяющими наступление известных формообразовательных реакций, а также в существовании взаимозависимостей между этими последними.
В процессах преобразования выражения мутаций происходит поэтому стабилизация процессов формообразования через развитие системы геномных и морфогенетических корреляций, связывающих организм в более устойчивое целое (И. Шмальгаузен, 1938а, 1942). При этом развитие нормы становится более защищенным не только от небольших мутаций, но и от внешних влияний. Руководящим фактором в этих процессах является стабилизирующая форма естественного отбора (т. е. постоянная элиминация неблагоприятных уклонений).
В. ИЗМЕНЕНИЯ ПРОЯВЛЕНИЯМУТАЦИЙ (ВОЗНИКНОВЕНИЕ ГЕТЕРОМОРФИЗМА И ПОЛИМОРФИЗМА)
Если мутация с изменчивым выражением колеблется между ясно видимым проявлением и полным его отсутствием, то говорят о неполном ее проявлении. Неполным может быть проявление полудоминантной мутации в гетерозиготном состоянии, а также рецессивной, а иногда даже полудоминантной мутации в гомозиготе. Неполное проявление указывает на особую лабильность данной мутации, на резко выраженную зависимость формообразования от наследственных свойств каждой особи и притом именно от ничтожных различий в генотипе, обусловленных присутствием тех или иных малых мутаций (модификаторов). Вместе с тем лабильность мутации выражается и в не менее резко выраженной зависимости формообразования от изменений в факторах внешней среды (температуры, влажности, корма и т. п.). Это служит основой развития генотипического и фенотипического разнообразия форм внутри популяции.
Так как изменчивость выражения и проявления указывает на значительную зависимость развития данных признаков (мутантов) от небольших генотипических различий, а также от изменений в факторах внешней среды, то этим самым дана возможность быстрого изменения этого выражения в результате естественного отбора. Если мутация неблагоприятна, то ее выражение должно бы исчезнуть в результате стабилизирующего отбора в пользу нормы. Если же она благоприятна, то она должна бы быть усилена до полной доминантности и войти в состав нормы в результате ведущей формы естественного отбора.
Если это не происходит с целым рядом особенно изменчивых мутаций, то для этого должны быть свои основания. Очевидно, в природной обстановке имеются условия, поддерживающие эту изменчивость. В пользу этого говорит и распространенность полудоминантных мутаций с неполным проявлением в природных популяциях (С. Гершензон, 1941а, б). При этом доминантность таких мутаций выражается сильнее при скрещиваниях внутри своей популяции, чем при скрещивании с другими популяциями (Р. Берг). Это означает отсутствие отбора на снижение их выражения в гетерозиготе, а быть может, даже свидетельствует о наличии положительного отбора. Можно думать, что существует система модификаторов, усиливающих выражение этих мутаций, и что эта система при скрещиваниях с другими популяциями частью разрушается.
Быть может, в таких мутациях их морфологическое выражение не столь неблагоприятно, как это нам кажется, и можно предполагать наличие прочной (т. е. неустранимой) корреляции с какими-либо положительными изменениями физиологического характера. В таком случае мы имеем лишь частично вредные (частично полезные) мутации.
Распространенность таких мутаций в больших популяциях заставляет также думать, что во многих случаях она относится к группе лишь условно вредных мутаций. Широкое распространение популяции, сталкивающейся с разнообразными микрогеографическими условиями, позволяет предполагать, что некоторые мутации могут оказаться в относительно благоприятном положении в определенных частных условиях существования (С. Гершензон) (или в известных сезонах, или при случайных изменениях в факторах среды). В этом случае концентрация мутации будет поддерживаться на известном уровне соответственно частоте встречаемости данных частных условий существования. Возможно, что как гетерозигота, так и обе гомозиготы в процессе эволюции разовьют свои условно наиболее благоприятные выражения. Возможно, наконец, и установление более сложных, дигибридных комбинаций со многими, вполне в известных условиях приспособленными формами.
Это приведет к установлению постоянного гетероморфизма данного вида. Частными случаями такого гетероморфизма являются различные окраски богомолов и различные формы мимикрии самок у Papilio polytes и P. dardanus. В других случаях в популяциях встречаются и безусловно вредные, но все же «доминантные» мутации. Как уже упоминалось, в таких случаях можно думать о доминантности как об особой форме «защиты» популяции от распространения вредной мутации. Такая «защита» могла (по Р. Л. Берг) развиться лишь в результате межгрупповой конкуренции, в которой побеждали более «здоровые» популяции. Однако мы видели, что мутации, по Фишеру, возникают как полудоминантные. Элиминируемые мутации не могут менять в процессе эволюции своего выражения и тогда продолжают возникать вновь в качестве таких же «доминантных» мутаций.
С другой стороны, мутации с изменчивым выражением и проявлением обнаруживают значительную зависимость не только от индивидуальных свойств генотипа, но и от изменений в факторах внешней среды. Их лабильность проявляется, следовательно, и в индивидуальных реакциях на те или иные частные условия (локальные, временные, сезонные). Такие реакции также могут оказаться частично благоприятными для организма. Они могут иметь хотя бы до известной степени, т. е. в некоторых случаях, положительное значение и могут быть преобразованы в процессе отбора, как показывают эксперименты М. Камшилова (1939б). В таких реакциях мы видим зародыши способности организма к индивидуальным адаптациям как физиологического, так и морфологического характера. Последние обозначаются как приспособительные модификации и являются источником более или менее ясно выраженного полиморфизма данного вида организмов.
Г. СТАБИЛИЗАЦИЯ БЛАГОПРИЯТНЫХ ВЫРАЖЕНИЙ МУТАЦИЙ
Если известные выражения отдельных мутаций имеют хотя бы в некоторых частных условиях существования положительное значение, увеличивая приспособленность организма или его общую жизненную стойкость, то эти выражения становятся предметом естественного отбора. Они преобразуются в процессах комбинирования и усиливаются до достижения известного оптимального уровня. Возможность усиления выражения мутации в гетерозиготном состоянии через ее комбинирование с другими мутациями доказана наблюдениями над дрозофилой. Рецессивная, связанная с полом мутация forked выражается у самца и у гомозиготной самки в укорочении щетинок на голове, груди и скутеллуме. Кроме того, щетинки изогнуты и приподняты. Гетерозиготная самка неотличима от дикой нормы. При ее комбинировании с другой рецессивной мутацией semiforked гетерозиготная самка forked получает, однако, явно промежуточное выражение, т. е. становится «доминантной» (точнее, полудоминантной; D. Lancefield, 1918, по R. Fisher, 1930). Конечно, такой значительный эффект лишь редко достигается комбинированием с одной мутацией, обладающей заметным модифицирующим влиянием. В естественном отборе на жизненную стойкость благоприятных мутаций материал доставляется многими малыми мутациями — модификаторами. Так как этот отбор происходит среди гетерозигот, то повышение положительного выражения мутации в гетерозиготе означает вместе с тем усиление ее доминирования (по меньшей мере в данном ее выражении). В этом случае возможно быстрое распространение мутации, переход части особей в гомозиготное состояние и отбор на такое же оптимальное выражение в гомозиготе, т. е. на полное доминирование и вхождение ее в состав новой, вполне стабильной нормы. Эволюция доминантности безусловно благоприятной мутации завершается, следовательно, вхождением нового признака или реакции в состав нормы. Вместе с тем в состав нормы включаются в качестве обязательных ее элементов и те модификаторы, которые подавляют неблагоприятные выражения мутации, а также те, которые выправляют и усиливают благоприятные выражения.
Таким образом, постепенно создается определенная генетически сбалансированная система (система геномных корреляций), которая не может быть нарушена без серьезных последствий для нормального формообразования. Устойчивость нормы не нарушается в результате этого постоянного включения в нее новых элементов. Наоборот, постепенное усложнение системы связей все более затрудняет изменение отдельных элементов этой системы. Если же такое изменение наступает, то оно является дезинтегрирующим и по своим последствиям может оказаться летальным. Этот путь эволюции мог бы завести в тупик, если бы не половой процесс и диплоидность (а также регуляторный аппарат), благодаря которым каждая мутация, возникая только в одном геноме, проходит в гетерозиготном состоянии предварительно долгий путь переработки под интегрирующим влиянием стабилизирующей формы естественного отбора.
Однако не всегда дело обстоит так просто. Мы видели, что условная вредность мутации (условная полезность) может привести к более или менее уравновешенному гетероморфизму. К гетероморфизму может привести и частичная вредность (частичная полезность) мутаций. Если благоприятное выражение мутации в результате естественного отбора усиливается в гетерозиготе и достигает оптимального уровня (и доминантности), а неблагоприятное выражение ослабляется и нейтрализуется (т. е. становится рецессивным) также лишь в гетерозиготе, то гомозиготизация данного изменения может вообще оказаться невозможной.
Во всех таких случаях гетерозигота окажется значительно более жизнеспособной, чем гомозигота. Это наблюдается иногда уже на отдельных мутациях. Так, в наблюдениях Леритье и Тессье (Teissier. L'Heritier, 1937) мутация ebony в гомозиготном состоянии не могла конкурировать с дикой нормой и вытеснялась ею, а в гетерозиготном состоянии она поддерживала свое существование на определенном уровне концентрации (20%). Здесь получен, следовательно, экспериментальный «равновесный» полиморфизм искусственной популяции. То же самое происходит, однако, и в природе, если какая-либо мутация оказывается полезной лишь в одном своем выражении. В особенности это касается мутаций с рецессивным летальным или стерильным влиянием. В этом случае гомозиготная мутация оказывается невозможной, а гетерозигота все время расщепляется с образованием гомозиготной нормы и гетерозиготной мутации. Популяция становится равновесно-диморфной.
Если гетерозигота обладает заметными преимуществами перед нормой, то это легко приводит к фиксированию какой-либо другой летали и в гомологичной хромосоме. Тогда приспособленная гетерозиготная форма становится на место нормы и дает начало только таким же гетерозиготным особям (обе гомозиготные формы летальны). Такая система «сбалансированных деталей» способствует, однако, и дальнейшему накоплению неблагоприятных (в гомозиготе) мутаций (Мёллер). Это — крайний случай. В более мягкой форме такие процессы оказываются весьма распространенными.
Примерами высокогетерозиготных популяций, насыщенных деталями, могут послужить виды божьих коровок и, в частности Adalia bipunclata. Гетерозиготность особей обнаруживается как в их разнообразной окраске, так и в физиологических признаках.
Вследствие большой насыщенности популяций Adalia bipunctata деталями, родственное разведение (путем скрещивания братьев с сестрами) оказывается чрезвычайно затрудненным. Оно сопровождается большой гибелью на всех стадиях развития (Я. Я. Лус, 1947)
Отбор идет в природных условиях по гетерозиготам, на базе отбора и фиксирования сбалансированной системы модификаторов, подавляющих вредные выражения мутаций и выдвигающих хотя бы условно благоприятные их выражения. Естественно поэтому, что именно гетерозигота должна, несмотря на ее лабильность, обладать большей приспособленностью и жизнеспособностью. Создаются, таким образом, сложные генетические системы, сбалансированные именно в гетерозиготном состоянии. Полная гомозиготизация таких систем вообще невозможна. Частичная же гомозиготизация, практически достигаемая путем инцухта, ведет к снижению жизнеспособности и к уменьшению плодовитости, как это нередко И наблюдается. Поэтому большая жизнеспособность гетерозиготы и ведет обычно к развитию механизмов, обеспечивающих скрещивание животных и поощряющих перекрестное опыление у растений (R. Fisher, 1930).
Устойчивость гетерозиготы означает, конечно, отсутствие неблагоприятных выражений наличных мутаций в их положительных выражениях. Такое изменение выражения мутаций создается путем комбинирования многих малых мутаций под руководящим влиянием стабилизирующего отбора, т. е. в данном случае отбора в пользу установившейся гетерозиготной нормы. Оно может вести и к тому, что гетерозиготная мутация окажется более устойчивой и жизнеспособной, чем каждая из гомозигот (т. е. как мутация, так и прежняя «норма»). Это явление сверхдоминирования является также результатом действия стабилизирующего отбора по гетерозиготам.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 1336; Нарушение авторских прав?; Мы поможем в написании вашей работы!