КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Возникновение приспособлений (адаптациогенез) 3 страница
|
|
|
|
Температурные реакции животных выражаются гораздо чаще в изменении пигментации, однако такие модификации имеют лишь редко явно адаптивный характер. В особенности у птиц известны закономерные изменения окраски (правило Глогера, по которому в теплых и влажных местностях интенсивность пигментации возрастает, а в холодных и сухих окраска ослабляется,— это правило подтверждалось Алленом, Гёрницем, П. Серебровским и Реншем). Отчасти, по крайней мере, такие изменения могут быть модификационными, как это показывают некоторые эксперименты. Адаптивность этих изменений, однако, сомнительна. Пигментообразование — сравнительно простой физиологический процесс, зависящий от течения ферментативных окислительных реакций. При их ускорении под влиянием повышенной температуры окислительные процессы идут дальше и ведут к образованию более темных пигментов (черных эвмеланинов и желто-красных феомеланинов). В таком случае некоторое побледнение окраски у животных холодных стран, а также у пустынных животных может быть отнесено частично и за счет модификаций, адаптивность которых является лишь побочным результатом физиологических изменений процесса пигментообразования.
Однако известны и такие изменения окраски, которые имеют явно приспособительный характер, согласованный с изменениями общего фона и освещения окружающей среды. Это касается куколок некоторых бабочек, некоторых рыб, амфибий и рептилий. В большинстве случаев масштаб этих изменений весьма незначителен. Однако у некоторых позвоночных способность изменять свою окраску так развита, что издавна обращала на себя внимание натуралистов. Она основывается главным образом на сокращении отростков и на перемещении хроматофоров, реже — на перемещении пигментных зерен в отростках хроматофоров и иногда — на усиленном их размножении. Только последнее, имеющее место у молодых саламандр, может рассматриваться как зависимый морфогенетический процесс. Характер зависимости, однако, не установлен. В других случаях изменение окраски оказывается легко обратимым процессом перемещения или сокращения хроматофоров, осуществляемым через нервную систему в результате зрительных восприятий животного. Сюда относятся известные защитные реакции камбалы, окрашивающейся под цвет песка или гравия, на котором она располагается, а также классический пример хамелеона, меняющего свою окраску с поразительной скоростью и достигающего очень значительного разнообразия цветов (от светло-серого до темно-зеленого). Ясно, что эти приспособительные изменения относятся к группе легко и быстро обратимых физиологических реакций, которые мы оставляем здесь без рассмотрения (так же как и всю большую область явлений приспособительных двигательных реакций организмов — рефлексов, инстинктов и поведения животных).
К категории физиогенных модификаций физиологического характера следует отнести и изменение количества эритроцитов, а также содержания гемоглобина в крови позвоночных животных, в зависимости от атмосферного давления. Заметное увеличений этих показателей установлено у ящериц и млекопитающих, в частности и у человека, при их перемещении в горные местности (или при экспериментах в барокамере). Эти изменения бесспорно адаптивны. Они устанавливаются довольно быстро и полностью обратимы. По-видимому, они определяются парциальным давлением кислорода в самой крови и соответственно представляют одно из широко распространенных явлений функциональных компенсаций, которые часто проявляются при недостаточности некоторых жизненных функций.
в. Функциональные модификации. Разделение модификаций на общефизиологические, физиогенные и функциональные имеет лишь весьма условный характер, так как все они определяются физиологическими процессами, т. е. основываются на жизненных функциях организма. Поэтому можно было бы и все модификации, так же как и все адаптивные физиологические реакции, обозначить как функциональные приспособления в широком смысле, или аккомодации. Для наших целей, однако, удобнее понятие аккомодации дифференцировать. В данном случае мы обращаем внимание на различную активность организма. Функциональными в узком смысле слова мы будем считать адаптации, возникающие в связи с изменениями активности самого организма.
Если физиогенные адаптации характерны главным образом для растений и осуществляются через посредство возникающих, еще не дифференцированных тканей, то функциональные адаптации свойственны в основном животным и осуществляются путем перестройки дифференцированных уже и нормально функционирующих тканей и органов. Они определяются всегда ролью известной ткани или органа в жизнедеятельности организма в целом (рис. 29).
Во всех таких адаптивных реакциях усиленная функциональная нагрузка ткани или органа ведет к гипертрофии, а систематическая недогрузка — к ослаблению органа и явлениям атрофии. К сожалению, физиологический механизм этих изменений не изучен в достаточной мере. Если нормальное функционирование органа связано с поддержанием известного равновесия между процессами диссимиляции и ассимиляции, то в состоянии покоя или при перенапряжении органа происходит, по-видимому, не столько нарушение количественной стороны этого равновесия, сколько изменение его качественной характеристики. При отсутствии специфических продуктов разрушения данного органа нет и восстановления его специфических элементов. Обмен веществ ограничивается одним лишь основным обменом. Обмен функционирующего органа имеет, очевидно, не только количественно, но и качественно иной характер. Специфические продукты обмена функционирующего органа являются, вероятно, стимуляторами процессов физиологической регенерации дифференцированных элементов органа. Увеличение концентрации этих продуктов выше известного уровня ведет к преобладанию восстановительных процессов над функциональным распадом. Функциональная гипертрофия приводит к установлению нового равновесия па более высоком уровне концентрации специфических продуктов обмена. Наиболее характерны явления функциональной гипертрофии в мышцах. Тренировка мышц ведет к их усилению вследствие увеличения толщины отдельных мускульных волокон. Сила мышцы, как известно, пропорциональна ее поперечному сечению. Сходны и явления функциональной гипертрофии в нервной ткани. И здесь они связаны с увеличением массы нервных клеток и увеличением поперечника их нервных отростков. Однако у зародыша или личинки ЕОЗМОЖНО и увеличение числа нервных клеток и, следовательно, и нервных волокон. Это видно из ряда экспериментальных исследований (над амфибиями).
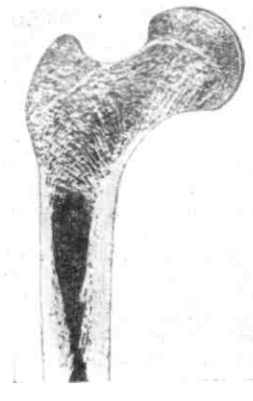
Рис. 30. Структура верхней части бедреннод кости человека. Костные пластинки располагаются в губчатом веществе кости по линия:» максимального натяжения и сжатия (Wolff, 1892)
В опорных тканях реакция на увеличение механической нагрузки выражается и в увеличении объема, и в перестройке самой структуры, если эта нагрузка меняет свое направление. Эти реакции наиболее изучены на волокнистой и эластической соединительных тканях и на костной ткани. Адаптивность изменений строения опорных тканей исключительно высока. В соединительных тканях она выражается в развитии обыкновенных и эластических волокон по линиям натяжения. В костной ткани она выражается в развитии более сложной внутренней структуры в виде системы костных пластинок, располагающихся главным образом по линиям сжатия и линиям растяжения. Там, где эти направления расходятся, образуются более сложные губчатые структуры с перекрещивающимися костными перекладинами (рис. 30).
Исключительно пластична кровеносная система. Она в высшей степени быстро перестраивается согласно функциональным требованиям возросшей массы тканей или их возросшей активности. Даже во взрослом организме легко устанавливается коллатеральное кровообращение при облитерации отдельных сосудов.
Такая же легкая приспособляемость обнаруживается и во многих внутренних органах. Известна высокая способность сердечной мышцы к тренировке РГ К компенсации при нарушениях нормального кровообращения. Точно так же возможны и адаптивные изменения дыхательной поверхности жабер у личинок земноводных (аксолотля) и дыхательной поверхности легких у наземных позвоночных. Весьма пластичны и почки позвоночных. Даже у млекопитающих возможны новообразование вторичных почечных канальцев у взрослого животного и далеко идущие явления компенсации при усиленной функциональной нагрузке, падающей на одну или обе почки.
Во всех этих функциональных адаптациях или компенсациях имеется нечто общее. Самое функционирование связано с процессами, ведущими к поддержанию данного органа, ткани или системы на известном уровне развития, и оно же ведет к превышению этого уровня при повышенной нагрузке. По всей вероятности, такого рода функциональные зависимости устанавливаются вместе с возникновением данной дифференцировки в процессах взаимной обусловленности исторического развития органа (ткани, системы) и его функции. Это историческое развитие всегда поддерживалось, с самого начала данной дифференцировки, взаимной обусловленностью индивидуального развития структуры органа (системы, ткани) и его функции. Можно думать, что и эволюция таких структур шла в значительной мере на базе этой взаимообусловленности структур и функций в индивидуальном формообразовании (см. дальше раздел Г. Значение адаптивных модификаций в эволюции. Стр. 305).
Иными словами, мы полагаем, что новые функциональные дифференцировки возникают всегда на базе жизнедеятельности самого организма. В связи с новым расчленением функций устанавливаются и новые структуры как результат функциональной деятельности организма. Функциональные дифференцировки возникают под влиянием самой функции, а в дальнейшей эволюции происходит их стабилизация и включение через смену морфогенных факторов в число автономно развивающихся структур. При таком понимании эволюции активных функциональных структур вполне понятно и не требует специальных пояснений то обстоятельство, что эти структуры фиксируются сразу же вместе с теми физиологическими механизмами (специфика обмена веществ), которые сопровождают их возникновение. Сопутствующие условия развития становятся в процессе эволюции необходимыми условиями реализации известных структур (см. стр. 54 о внешних условиях развития). Продукты специфического обмена получают при этом значение морфогенных веществ, стимулирующих развитие именно данных структур.
К функциональным адаптациям можно отнести также образование мозолей, т. е. утолщений рогового слоя кожи, наблюдаемых у птиц и особенно у млекопитающих, в тех частях общих покровов, которые испытывают постоянное давление или трение. Такие реакции можно рассматривать также как явления функциональной гипертрофии, основанной на стимуляции процессов физиологической регенерации.
Функциональным адаптациям мы приписываем очень большое значение в возникновении новых дифференцировок в процессе эволюции высших позвоночных. Как это осуществляется, мы рассмотрим несколько далее. Однако отметим тут же, что это понимание генезиса функциональных дифференцировок подводит нас в то же время и несколько ближе к пониманию самого возникновения способности к функциональным модификациям.
г. Индивидуальная коадаптация органов. Индивидуальная приспособляемость организма выражается в физиологических и морфологических изменениях, связанных с изменениями в факторах внешней среды. Собственно физиологических реакций мы здесь не касаемся, как бы велики ни были их адаптивности и их значение в эволюции. Что же касается морфологических адаптации, выражающихся в более стойких изменениях формы и структуры организма и его органов, они, как сказано, могут иметь зависящую от внешних факторов общефизиологическую основу (явления роста и размножения). Они также могут строиться на частных зависимостях формообразовательных процессов от внешних факторов (температуры, света, влажности) или основываться на их зависимости от функциональной нагрузки (диктуемой, очевидно, положением организма во внешней среде).
Части лабильного организма и его органы приспособляются, однако, не только к внешней среде. Можно говорить и о взаимном приспособлении органов внутри организма. И в этом случае речь может идти о взаимозависимостях в общефизиологических процессах роста, ведущих к изменениям в пространственных соотношениях, о взаимозависимостях в специфических факторах морфогенеза (морфогенетические корреляции) или о взаимозависимостях в функциях (эргонтические корреляции). Все эти связи могут вести к согласованным преобразованиям в соотношениях органов и их частей (если они имеют регуляторный характер).
Наличие таких взаимозависимостей различного рода представляет эмпирический факт. Мы различаем геномные, морфогенети-ческие и эргонтические корреляции (Шмальгаузен, 1938а, 1939а, 1942). Из них часть морфогенетических и, по-видимому, все эргон-тические корреляции имеют регуляторный характер. Это означает, что изменение одного звена (зачатка, ткани, органа) влечет за собой согласованное изменение другого звена (зачатка, ткани, органа) корреляционной цепи. Морфогенетические и эргонтические корреляции регуляторного характера строятся на базе элементарных морфогенетических взаимозависимостей, проявляющихся уже в отдельных мутациях (выражения множественного и многостепенного плейотропизма см. далее гл. 4). На отдельных мутациях и на морфозах нередко уже ясно видны взаимозависимости в изменении соотношения органов. Они изучались в последнее время Рапопортом (1941) на хемоморфозах и М. Камшиловым, а также Р. Берг на мутациях с изменчивым выражением. М. Камшилов установил существование зависимостей между различными выражениями одной мутации. Путем искусственного отбора, а также при разных условиях развития достигался сдвиг в виде закономерного ряда последовательных изменений различных частей (органов). М. Камшилов видит в этом указание на существование взаимозависимостей типа градиентов. Р. Берг (1943) пришла к такому же выводу. И. Рапопорт предпочитает говорить о морфогенетических взаимозависимостях в более общей форме.
Во всяком случае, эти исследования показывают, что в мутациях обнаруживается наличие корреляций и, соответственно, в них происходит взаимозависимая перестройка частей. Такая же взаимозависимость обнаруживается и в морфозах. В процессе эволюции такие элементарные взаимозависимости приобретают целесообразно-регуляторный характер. Как уже сказано, многие морфогенетические взаимозависимости (корреляции) в развитии частей нормального организма (как они изучаются «механикой развития») имеют именно регуляторный характер. Такой же характер имеют, по-видимому, и все эргонтические корреляции. Эргонтические корреляции основываются на тех же реакциях, что и функциональные адаптации. Механизм адаптации и коадап-тации органов здесь по существу совпадает. Разница лишь в источнике изменения — лежит ли он во внешней среде или в изменении другого органа. Если увеличение количества эритроцитов и гемоглобина, увеличение сердечной мышцы или увеличение дыхательной поверхности легких определяются уменьшением парциального давления кислорода в крови, то по сути совершенно безразлично, что является первоисточником этих изменений — пониженное атмосферное давление в горной местности или повышенная активность организма в связи с трудностью добычи необходимого питания или при переходе к иному способу движения, требующему более значительной работы мышц (например, при летании). Если увеличение числа почечных канальцев определяется повышенной концентрацией продуктов обмена в крови, то опять-таки механизм этого изменения один и тот же как в том случае, если это увеличение зависит от изменения качества пищи или от изменения активности организма и количества пищи, так и в том случае, если оно зависит от более глубокого изменения самого обмена или от простого увеличения массы самого животного.
Практически явления адаптации организма к внешней среде и явления коадаптации органов неразграничимы, так как любая адаптация известных частей к изменению в факторах среды неизбежно сопровождается и соответственной коадаптацией, т. е. взаимозависимым изменением других частей и, строго говоря, всей организации. Точно так же и наоборот: любая перестройка организации меняет положение организма во внешней среде и изменяет приспособления отдельных частей к различным условиям этой среды.
Наличие системы корреляций регуляторного характера (об ее происхождении см. главу 4 этого раздела) приводит к тому, что изменение в процессе эволюции (или даже в результате индивидуальной адаптации) одной части или органа влечет за собой согласованное изменение другой части или органа. Такое согласование происходит отчасти именно в процессе индивидуального развития. Взаимное приспособление органов достигается путем согласования пространственных соотношений через морфогене-тические взаимозависимости, коррелятивный рост и функциональные взаимозависимости. Благодаря этому приспособления организма к окружающей среде приобретают целостный характер. Индивидуальная адаптация известных органов к измененным факторам внешней среды дополняется такой же коадаптацией других органов, так что в результате происходит согласованная перестройка всей организации. Можно говорить об установлении новой адаптивной нормы.
Однако система морфогенетических корреляций регуляторного характера достигает большой сложности только у позвоночных животных. Поэтому адаптации растений и большинства животных далеки от такой совершенной внутренней согласованности — они ограничиваются нередко частными изменениями отдельных органов. Впрочем, для организмов, живущих на грани различных сред и попадающих в весьма различные условия существования, даже физиогенные адаптации могут (главным образом через систему геномных корреляций) приобрести целостный характер вполне выработанных «адаптивных норм».
В. АДАПТИВНЫЕ НОРМЫ И ИХ СМЕНА
В организмах со слабым развитием системы регуляторных корреляций коадаптация частей осуществляется лишь медленно, в порядке естественного отбора наиболее жизненных, наиболее гармонических соотношений.
Изменение отдельной части или признака организма соответственно изменению фактора внешней среды нарушает до известной степени гармоническую согласованность частей. Этим самым значительно снижается жизненная и селекционная ценность любого положительного изменения форм реагирования. Поэтому способность к частным адаптациям дополняется по меньшей мере и другими реакциями, т. е. некоторыми зависимостями, позволяющими внести большую согласованность в изменения всего организма в целом. Такие зависимости имеют на первых ступенях своего развития характер геномных корреляций. В процессе эволюции вырабатываются, таким образом, типичные реакции, ставящие весь организм в новое положение. Между типичными формами приспособления образуются разрывы, и переход от одной типичной формы к другой приобретает характер скачка. В особенности это имеет место при резком изменении положения во внешней среде и доминирующем значении физиогенных адаптации. Поэтому такие переходы от одного типа приспособления к другому более ярко выражены у растений, живущих на грани различных биотопов и попадающих систематически и случайно то в один, то в другой биотоп. При развитии типических норм реагирования организма, в виде различных типов приспособления, особое значение приобретают регуляторные процессы.
При небольшой интенсивности внешнего фактора может не наступать никакой видимой реакции. На известном уровне этой интенсивности, соответствующем нижнему порогу реактивности организма, наступает сразу в полном своем выражении реакция, зависимая от данного фактора. Дальнейшее изменение интенсивности не изменяет результата формообразовательных процессов, вплоть до некоторого уровня, соответствующего верхнему порогу реактивности, по достижении которого данная реакция вновь выпадает, заменяется морфозом или совершенно иной адаптивной реакцией. В особенности интересны, конечно, случаи полного переключения одной реакции на совершенно иную, но также приспособительную реакцию.
Между нижним и верхним порогами нормальной реактивности организма, выражающейся в реализации адаптивной модификации, лежит широкий диапазон возможных изменений интенсивности, на которые организм как будто вовсе не реагирует. Это указывает на частичную утрату детерминирующего значения внешнего фактора. Последний выступает в роли стимулятора, лишь возбуждающего известную цепь формообразовательных процессов, которые протекают в основном под влиянием внутренних факторов развития. Повышение интенсивности внешнего фактора за пределы нижнего порога реактивности организма не оказывает заметного влияния на течение этих процессов вследствие очевидного наличия регуляторных механизмов, противодействующих этим влияниям. Такую форму индивидуального развития, зависящего в своем осуществлении от известного минимума интенсивности внешнего фактора, который дает лишь первый толчок к развертыванию внутреннего механизма типично детерминированного формообразовательного процесса, мы называем авторегуляторным развитием.
Процессы авторегуляции представляют адаптивные ограничения нормальной реактивности известными пределами. С таким ограничением реактивности известными пределами «нормы» мы познакомились уже при рассмотрении общефизиологических реакций (в явлениях роста). Возникновение подобных регуляторных механизмов на базе элементарных реакций, свойственных отдельным мутациям, нетрудно себе представить, так как такие реакции практически всегда укладываются в известные рамки, которые могут сдвигаться как в одну, так и в другую сторону при комбинировании разных мутаций и в процессе естественного отбора различных комбинаций.
Типичная окраска горностаевого кролика (рис. 8) — белый с черными носом, ушами, хвостиком и концами лапок — развивается, по Н. Ильину, нормально при температуре среды от 2 до 14°. При температуре выше 14–16° концы лапок становятся белыми. В пределах от 2 до 14° окраска не зависит от изменений температуры среды. Необходим минимум в 2° для того, чтобы типичная окраска горностаевого кролика проявилась в полной мере; 2° являются нижним пороговым уровнем нормальной окраски, а 14°— верхним порогом нормальной реактивности горностаевого кролика на температурные изменения. При более высокой температуре, как сказано, развиваются белые лапки. В пределах от 16 до 25° мы имеем также вполне типичную и устойчивую вторую «норму» —- белый кролик с черными ушами, носом и хвостиком. При температуре выше 25° начинается побеление ушей, а при 29° начинают белеть нос и хвостик. Такая высота различных порогов реактивности кожи горностаевого кролика характерна, однако, только для московской расы. У западных рас температурные пороги находятся на гораздо более высоком уровне (для кожи бока нижний порог белой окраски у московской расы + 2°, у польской + 11°, а у саксонской +21°). Имеются и индивидуальные наследственные различия пороговых уровней у отдельных кроликов.
Все это показывает возможность смещения пороговых уровней реактивности как в индивидуальных уклонениях (мутациях) и их комбинациях, так и в результате естественного отбора, ведущего к образованию различных наследственных форм.
Таким образом, подобные реакции вводятся в ограниченное русло более определенной «нормы», приспособленной именно к известным условиям внешней среды (например, к известным пределам температуры). При этом для разных условий могут вырабатываться и характерные для них и одинаково приспособленные различные «нормы» (для низких зимних температур одна норма окраски, например белая; для более высоких, летних температур — другая норма, например серая или бурая).
Известны многочисленные случаи сезонного полиморфизма с реакциями именно такого рода. Если куколка Araschnia (Vanessa) levana зимует или в эксперименте подвергается любым температурам ниже 0°, то развивается типичная рыжая весенняя форма А. levana. При температурах, превышающих + 2°, развивается типичная темная, летняя форма A. prorsa.
У животных адаптивный полиморфизм имеет именно почти всегда сезонный характер. В особенности характерны явления «цикломорфоза» у планктонных организмов — коловраток и дафний. Здесь формообразование зависит в своей специфике от взаимосвязанного комплекса изменений температуры и питания и выражает приспособление к парению в воде различного удельного веса (увеличение поверхности у летних форм).
Можно привести многочисленные примеры модификационного полиморфизма, связанного с существованием регуляторных механизмов, определяющих развитие нескольких приспособительных норм у растений. Как раз у растений, лишенных возможности активно перемещаться и переходить из одного биотопа в иной, более подходящий, большое значение имеет способность к физиогенным адаптивным модификациям и, в частности, нередко имеются хорошо дифференцированные целостные приспособительные нормы. Как уже упомянуто, это в особенности характерно для растений, живущих на грани резко различающихся биотопов. В этом случае регуляторный характер реакции, полностью реализующейся в типичном виде на известном уровне интенсивности внешнего фактора, выступает вполне ясно.
У водяной гречихи (Polygonum amphibium) при известной степени влажности (даже без непосредственного воздействия воды) внутренний механизм развития воздушных листьев сразу переключается на механизм развития водных «плавающих» листьев (Goebel, 1928). У стрелолиста (Sagittaria sagittifolia) при известном ослаблении освещения реакция образования наземных стоячих листьев переключается на развитие водных лентовидных листьев (рис. 31). Более сложный комплекс факторов действует при модификациях растений в горных местностях. Здесь при комбинированном влиянии интенсивной инсоляции и низкой температуры развивается характерный альпийский габитус (низкий стебель с прикорневой системой обычно опушенных листьев, глубокие корни). В менее яркой форме такая множественность реакций свойственна очень многим растениям, встречающимся на различных почвах и вообще в различных экологических условиях («эко-фены» Турессона).

Рис. 31. Изменение формы листьев на одном и том же экземпляре стрелолиста Sagittaria sagittifolia в зависимости от условий среды. Пример нескольких типичных адаптивных модификаций (норм). По Курсанову
Во всех этих случаях мы имеем явления модификационного полиморфизма, связанного с авторегуляторным типом развития. Каждая форма вполне приспособлена к известному комплексу внешних условий, реально встречавшихся и встречающихся в жизни данного вида в качестве случайных, периодических (сезонных) или локальных явлений. Вид в целом приспособлен не в одной своей форме, а в двух или нескольких формах, и не к одному сезону или биотопу с характерным для него комплексом условий, а к нескольким. Поэтому, в случае изменения среды, связанного с выпадением известного комплекса условий, характеризующего один из сезонов или один из биотопов, к которым вид был приспособлен, вид не вымирает, так как остается еще иной комплекс факторов, к которому данный вид был уже приспособлен. Заболачивание почвы, распространение леса или, наоборот, отступаний леса и развитие степи приводят к обязательной реализации тех реакций, которые раньше осуществлялись лишь случайно, периодически или локально (водная форма, теневая, ксерофитная и т. п.). Происходит «смена адаптивных норм», при которой прежняя главная реакция теряет свое значение и уступает место новой, бывшей раньше лишь второстепенной. Все реакции приобрели уже заранее целостный характер, и это обеспечивает немедленную перестройку всего организма даже при быстрых изменениях физических факторов внешней среды.
Прибрежное или болотное растение (рис. 32), иногда заливаемое водой и приобретшее уже способность развивать водные, листья, может в любое время перейти к чисто водной жизни. Растение континентального климата, теряющее иногда при засухе свои листья и приобретшее способность в этих случаях ассимилировать при помощи поверхностных тканей стеблей или листовых черешков, может при развившейся сухости климата быстро превратиться в типичного ксерофита, ассимилирующего при помощи стеблей и черешков (филлокладиев и филлодиев).
Эти представления о смене адаптивных норм можно было бы развить и дальше. Можно говорить о выделении новых норм, об увеличении числа норм, о расширении зоны адаптивности, о замене главной нормы второстепенной, о сокращении числа норм и сужении зоны адаптивности реакций (специализация). Однако значение самого принципа здесь освещено в достаточной мере, а детализация завела бы нас далеко за пределы разбираемой проблемы «факторов» эволюции.
Г. ЗНАЧЕНИЕ АДАПТИВНЫХ МОДИФИКАЦИЙ В ЭВОЛЮЦИИ
Вопрос о значении адаптивных модификаций в эволюции был в свое время правильно оценен Ч. Дарвином, хотя позднейшие «дарвинисты» видели в его высказываниях по этому поводу лишь ненужные «уступки» ламаркизму. Только у Л. Моргана, Болдуина и ряда других дарвинистов мы находим превосходную, до сих пор непревзойденную разработку этого вопроса с дарвинистических (в общем) позиций. К сожалению, позднейшие успехи генетики привели к распространению неодарвинистических представлений. Адаптивным модификациям при этом не отводилось вообще какого-либо места среди факторов эволюции. Только в новейшее время этот вопрос был вновь поставлен рядом советских авторов (Кирпичников, 1935, 1940; Лукин, 1935, 1936, 1939; Шмальгаузен, 1938а, 1939а, 1940а, 1941), которые рассматривали его с различных точек зрения. Однако и до сих пор вопрос о значении адаптивных модификаций не получил всестороннего освещения и, главное, не получил еще достаточной оценки. Я попытаюсь отметить здесь хотя бы основные положения для будущей его разработки. На первом месте я бы поставил значение самой лабильности, т. е. способности к адаптивным модификациям (более широкого вопроса об индивидуальной приспособляемости в физиологических реакциях и нервной деятельности я здесь не касаюсь) для жизни вида и для его потенциальной способности к эволюции. Затем я хочу отметить ведущее значение конкретных модификаций при изменении факторов внешней среды и при активном переходе в иную среду. Наконец, я не могу не обратить еще раз (Шмальгаузен, 1939а) внимания на значение способности к адаптивным функциональным модификациям в процессе установления действительно совершенно новых дифференцировок.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 688; Нарушение авторских прав?; Мы поможем в написании вашей работы!