КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Доминантность. Баланс нормального генотипа обычно достаточно защищен от нарушений со стороны небольших мутаций. В гетерозиготе это выражаетсяв доминировании нормы. 2 страница
|
|
|
|
Это усложнение ведет и к дальнейшему увеличению стойкости основ организации вследствие установления комплексного характера всех наиболее ответственных морфогенетических процессов и вследствие приобретения ими все более ясно выраженного ре-гуляторного характера. Путь развития таких регуляций, очевидно, сходен с рассмотренными уже процессами установления авторегуляции зависимых (от внешних факторов) форм индивидуального развития (см. I—5 и III—3, В).
Первоначальное действие «геногормонов» или индуцирующих веществ на известные ткани можно себе представить как влияние, нарастающее в своей интенсивности пропорционально концентрации веществ, доходящих до этих тканей (конечно, в известных пределах — от минимума, оказывающего вообще какое-либо влияние, до максимума, дающего наиболее значительную реакцию).
В процессе эволюции, вместе с изменением всего генотипа (и процессов внутриклеточного обмена) изменяются и качество и концентрация морфогенных веществ («активность» генов), а также и реактивность отдельных тканей. Концентрация морфогенных веществ, доходящих до реагирующих тканей, поднимается до уровня, обеспечивающего наступление реакции не только при уклонениях в факторах внешней среды (меняющих их продукцию или реактивность тканей), но и при мутациях. Это последнее достигается удвоением «нормальной» концентрации, дающей полный эффект в гомозиготе. Тогда проявляется доминирование нормы, т. е. нормальная концентрация морфогенных веществ и нормальное формообразование уже у гетерозиготы. Это означает вместе с тем и ограничение реактивности тканей, которые дают одну и ту же реакцию при простой (А) и при двойной (2А) дозе раздражителя. Однако новая мутация может означать не только уменьшение концентрации более чем вдвое (поэтому новые мутации часто полудоминантны), но и качественное изменение веществ. Она может означать и изменение реактивности тканей.
Поэтому нормальная реактивность тканей «охраняется» также известным «запасом», лежащим за пределами обычных колебаний концентрации морфогенных веществ (раздвигание обоих порогов реактивности). Кроме того, она «защищается» расширением этой реактивности и от качественных изменений тех же веществ (снижение специфичности индуцирующих веществ) и, наконец, «защищается» самой сложностью всего комплекса взаимодействующих факторов (зависимостью реактивности тканей и специфики морфогенных веществ от всего генотипа в целом и сложностью морфогенетических взаимозависимостей).
а. Раздвигание порогов нормальной реактивности тканей. Ес-тественый отбор наиболее жизнеспособных особей означает выработку оптимальной реактивности тканей при существующих обычных условиях поступления морфогенных веществ.
Последнее может подлежать некоторой изменчивости, зависящей как от мутаций, так и от уклонений в условиях развития. Весьма важно, чтобы оптимальная формообразовательная реакция всегда осуществлялась независимо от этих уклонений. Это и достигается в процессе эволюции путем стабилизирующего отбора (т. е. через элиминацию особей с уклонениями от нормальной формообразовательной реакции). Морфюгенетический эффект оказывается нормальным, почти независимо от концентрации морфогенных или индуцирующих веществ, по крайней мере в известных пределах обычно встречающихся уклонений от «нормальной» концентрации. Это ясно видно по материалам, изученным Гольд-шмидтом, в особенности по интерсексам непарного шелкопряда, а также и на примерах развития зависимых вторичнополовых признаков позвоночных.
Вырабатывается нижний, а быть может, и верхний порог концентрации веществ, необходимой для наступления нормальной (т. е. оптимальной) реакции (рис. 40). Чем шире раздвинуты оба порога, тем более развитие данной части оказывается обеспеченным авторегуляцией, как можно назвать это явление. Нормальное развитие обеспечено тогда и при мутациях, меняющих интенсивность образования морфогенных веществ в известных, конечно, пределах (особенно в сторону увеличения).
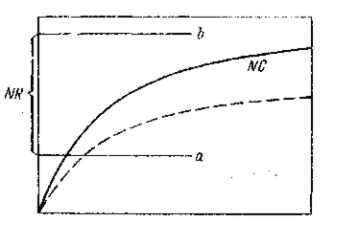
Рис. 40. Проявление известной нормы реактивности ткани. Развитие нормы стабилизируется через установление нижнего и верхнего порогов концентрации веществ, необходимой для наступления нормальной реакции. Морфогенные вещества теряют при этом свое детерминирующее значение и становятся в основном активирующими или освобождающими факторами. На оси абсцисс показано течение времени, а на ординате — нарастание концентрации веществ а — нижний и b — верхний порог нормальной реактивности ткани (NR)\ NC — кривая нормального нарастания концентрации морфогенных веществ. Прерывистой линией показано значительное снижение продукции морфогенных веществ, которая остается, однако, в пределах нормальной реактивности ткани. Такое изменение (мутация) имеет скрытые формы, но приближает организм к нижнему порогу нормальной реактивности и, следовательно, снижает устойчивость нормы, делает ее более доступной для выявления следующей мутации (или модификации), лежащей в том же направлении
Однако в действительности индуцирующие вещества поступают иногда даже не из одного источника, а из нескольких, и тогда нормальное развитие оказывается еще более обеспеченным. В этом случае возможны гораздо более значительные изменения в отдельных индукторах без того, чтобы нормальная комплексная реакция потерпела существенный ущерб. Получается нечто вроде «множественного обеспечения» нормального развития. Однако всякое уклонение, выходящее за пределы нижнего порога, приведет все же к выпадению известной реакции и явлениям недоразвития, а уклонение, выходящее за верхние пределы, может привести к заметному уклонению от нормального развития затронутых частей. Такие крайние уклонения и выявятся сразу в форме видимых «больших» мутаций или же в виде летали, если они затрагивают жизненно важный морфогенетический процесс.
В более сложных индукционных системах, связанных целой сетью многостепенных морфогенетических корреляций, каждое звено которых зависит от многих факторов типа активаторов или гормонов, достигается в конце концов относительно очень высокая степень устойчивости развивающейся авторегуляторной системы. В каждом звене имеется соотношение, которое может быть пояснено приведенной диаграммой (рис. 41), показывающей простейшую форму взаимодействия. Взаимные сдвиги во времени созревания активатора (A) и реактора (R) не оказывают влияния на нормальное течение морфогенетической реакции, пока они оба лежат в пределах между начальным и конечным пороговыми уровнями. Однако более значительное расхождение обоих компонентов индукционной системы, когда зрелая уже ткань активатора вступает в контакт с еще не созревшей или уже перезревшей и поэтому неактивной тканью (R), приводит сразу к полному выпадению данной морфогенетической реакции, а также и всех дальнейших зависящих от нее реакций. В результате происходит либо редукция этой части, либо очень большое недоразвитие, имеющее почти всегда летальные последствия, в особенности если нарушение коснулось одного из ранних и основных морфогенети-ческих процессов или жизненно важных органов.
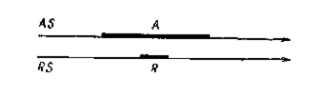
рис. 41. Упрощенная схема одного звена индукционной системы, состоящего из активатора (А) и реактора (R), которые изменяются во времени в направлении, показанном стрелками. Периоды зрелости обоих компонентов отмечены жирными участками линий. Система имеет регуляторный характер: она допускает в известных пределах возможность взаимных смещений во времени созревания взаимодействующих компонентов без того, чтобы реакция потерпела какой-либо ущерб.
То же самое касается и изменений в продукции активирующих веществ. Изменения, не выходящие за пределы нижнего и верхнего порогов концентрации веществ, вызывающей нормальную реакцию, не дадут никакого эффекта. При выходе же за эти пределы реакция либо вовсе не наступит (при недостаточной концентрации), либо, быть может, в некоторых случаях даст резкие уклонения от нормы (при чрезмерной концентрации). И в том и в другом случае процессы развития окажутся глубоко нарушенными и обнаружатся в виде явной мутации.
Огромное большинство мутаций практически не затронет ранних стадий развития частей, связанных системой морфогенетиче-ских корреляций. Обычные небольшие мутации могут получить видимое выражение лишь на поздних стадиях развития, когда система морфогенетических корреляций уже потеряла свое значение вследствие завершения основных процессов морфогенеза.
б. Снижение специфичности морфогенных веществ. Раздвигание обоих порогов нормальной реактивности ткани означает вместе с тем довольно широкую возможность уклонений от «нормальной» концентрации морфогенных веществ без нарушения нормального течения формообразовательного процесса. Количественные изменения морфогенных факторов в известных пределах остаются без влияния на развитие нормы. Однако то же самое устанавливется затем и но отношению к качественным изменениям. В процессах мутирования, несомненно, происходят и качественные изменения специфики морфогенных веществ, а не только количественные изменения в скорости их образования и в достигаемой концентрации. Элиминация возникающих при этом отклонений от нормы означает отбор особей, наименее реагирующих на качественные изменения морфогенных веществ. Таким образом, стабилизирующий отбор приводит к освобождению реагирующих тканей от влияния случайных изменений в концентрации и в специфических свойствах индуцирующих и вообще формативных субстанций.
Следовательно, параллельно с выработкой определенной нормальной реактивности тканей, в процессе эволюции теряют свое значение не только концентрация (в известных пределах, определяемых обоими порогами реактивности), но и специфические свойства морфогенных веществ (очевидно, также в известных пределах).
Специфика реакции определяется свойствами (генотипом) реагирующей ткани на данной стадии и в данном физиологическом положении, а значение морфогенных веществ низводится до уровня несиецифических активаторов.
Мы видели, что при авторегуляторном типе индивидуального развития факторы внешней среды теряют свое детерминирующее значение и становятся лишь раздражителями, освобождающими автономное в дальнейшем течение морфогенетичеоких процессов. Точно так же мы видим, что при прогрессивной автономизации онтогенеза внутренние морфогенные факторы теряют свою специфику и их значение снижается, наконец, до уровня раздражителей, освобождающих автономное течение специфических формообразовательных процессов («самодифференцирования»).
Все без исключения опыты обменной трансплантации зачатков, переносимых с зародышей одного вида на другой, показывают, что специфика морфогенной реакции определяется свойствами самой реагирующей ткани. Последняя образует типичные для нее дифференцировки даже и в том случае, если детерминирующее влияние исходит от индуктора, относящегося к другому виду, в котором такие дифференцировки никогда не осуществляются.
Уяче первые опыты обменной трансплантации эктодермы молодых гаструл разных видов тритона показали, что нервная трубка, развивающаяся из трансплантированной эктодермы, взятой с бока или брюха Triton cristatus, под влиянием крыши первичной кишки зародыша Т. taeniatus дифференцируется все же в точности по типу Т. cristatus. С другой стороны, эктодерма Т. taeniatus, которая, оставаясь на месте, дала бы начало обыкновенному эпидермису бока, дает после пересадки в область спины зародыша Т. cristatus под влиянием примыкающего снизу индуктора, т. е. крыши первичной кишки хозяина (Т. cristatus), начало нервной трубке. Однако эта последняя, хотя и образовалась под действием морфогенных веществ Т. cristatus, развивает все специфические особенности строения нервной системы Т. taeniatus. Подобных опытов было сделано очень много с индукцией хрусталика и других органов под влиянием чуждых индукторов. При этом были получены еще более удивительные результаты.
При трансплантации головной эктодермы зародыша аксолотля на бок головы зародыша тритона в этой эктодерме никогда не развиваются характерные для тритона опорные нити — «балансеры». Однако в обратном эксперименте пересадки туловищной эктодермы тритона на боковую часть головы зародыша аксолотля, под индуцирующим влиянием аксолотля (у которого балансеров никогда не бывает), в этой эктодерме развивались балансеры типичного для тритона строения (Harrison, 1925; Mangold, 1931). Далее производились обменные трансплантации головной эктодермы между тритонами, с одной стороны, и бесхвостыми амфибиями— с другой (Rotmann, 1935; Spemann и Schotte, 1932; Spe-raann, 1936). В эктодерме тритона развивались у зародышей жерлянок и других бесхвостых, под их индуцирующим влиянием, типичные тритоньи балансеры, а в пересаженной эктодерме бесхвостых развивались на тритонах, под их индуцирующим влиянием, типичные присоски и даже роговые челюсти головастиков.
Это показывает с полной ясностью, что видовая специфика зависимых морфологических структур определяется исключительно реагирующей тканью. Морфогенные влияния, хотя и необходимы в качестве факторов, определяющих осуществление, а также время и место наступления реакции, не определяют, однако, ее качества. Эти влияния лишены видовой специфичности.
Дальнейшие попытки проникнуть в природу морфогенных или индуцирующих веществ показали, кроме того, широкую замещае-мость одних индукторов другими и выявили возможность их замены другими тканями, в которых нельзя было и подозревать существования морфогенной активности, а также различными препаратами, в том числе и синтетическими химическими соединениями. Нервная пластинка, нормально развивающаяся под индуцирующим влиянием крыши первичной кишки (хордомезо-дермы), может быть индуцирована также всевозможными другими тканями и притом не только свежими, но и убитыми нагреванием, высушиванием или замораживанием (Holtfreter, 1932!, 1933). Оказалось даже, что некоторые ткани (эктодерма), в живбм состоянии не обладающие индуцирующими свойствами, приобретают таковые после высушивания при 60° (Holtfreter, 1933). Наконец, положительные результаты были получены с эфирными экстрактами зародышевых тканей (Waddington, Needham, 1934), с нуклео-протеидами и даже с синтетическими масляными кислотами и другими соединениями (Fischer, 1935 и др.)- Положительные результаты были получены и в опытах индукции хрусталика глаза при помощи различных тканей.
Опытным путем была получена индукция добавочных конечностей у тритонов путем пересадки в бок зародыша слухового пузырька (Balinsky, 1925). Позднее удавалось достичь того же и пересадкой обонятельных мешков, регенерационной бластемы и др. Н. Драгомиров (Dragomirov, 1929) показал, что волокнистое ядро хрусталика, нормально индуцируемое ретинальным листком глазного бокала, может быть в эксперименте индуцировано и слуховым пятном лабиринта или слуховым ганглием. Он же нашел (1936), что таким же мало специфическим влиянием контакта со слуховым пузырьком может быть индуцировано и развитие добавочной ретины в пигментном листке глазного бокала.
Следовательно, индуцирующие ткани и вещества не только лишены видовой специфичности и могут быть широко заменены соответствующими тканями и органами, взятыми от других животных, но не имеют и морфогенной специфики. В наибольшей степени это касается «первичного организатора», под влиянием которого развиваются нервная система и все осевые органы. В широкой мере неспецифичны и те влияния, которые вызывают образование добавочных конечностей. Однако здесь возможность индукции все же ограничена известной областью бока зародыша,— очевидно, здесь имеются необходимые добавочные влияния, быть может не лишенные некоторой специфичности. Кроме того, можно указать, что наиболее активным из чуждых индукторов является обонятельный мешок и именно его каналовая часть; несколько менее активен слуховой пузырек. Другие органы, очевидно, лишены этой активности. В индукции волокнистой структуры хрусталика нормальное влияние ретины может быть заменено весьма сходным, очевидно, влиянием слухового пятна или слухового ганглия, но оно не может быть заменено влиянием обонятельного мешка. Последний в этом случае оказывается недеятельным.
Таким образом, отсутствие органогенной специфики сказывается особенно ярко в индукторах наиболее общих и важных образований (осевых органов, конечностей) и гораздо менее выражено в частичной замещаемости индукторов частных структур (хрусталика и ретины глаза). Очевидно, это связано с тем, что формообразовательный аппарат более общих основ организации проделал уже более длинный путь эволюции в направлении утраты специфики морфогенетических влияний и установления максимальной автономности развития важнейших систем органов. Этим самым достигнута максимальная защита формообразования от каких-либо нарушающих влияний. Случайные изменения концентрации или химических свойств морфогенных веществ не могут тогда нарушить нормального развития основных органов животного. Это не значит, что сама «индукция» потеряла свое значение. Контакт индуктора и реактора определяет в индукционной системе если не специфику, то время наступления и, в особенности, согласованность положения и протяженность взаимозависимых морфогенных реакций и тем самым определяет возможность регуляции и сохранения гармонических соотношений между органами.
Эволюция онтогенеза идет в общем по пути прогрессивной автономизации важнейших морфогенных процессов, связанной с указанной уже регуляторной организацией индукционных систем. Эта организация допускает, кроме изменения в концентрации и в составе морфогенных веществ, также возможность более или менее значительных сдвигов во времени и в пространстве без нарушения нормального формообразования. Она имеет и в целом и в отдельных своих компонентах регуляторный характер.
Любой индифферентный еще зачаток допускает весьма далеко идущие экспериментальные вмешательства без нарушения дальнейшего формообразования. Если зачаток действительно совершенно индифферентен, то вполне понятно, что и часть зачатка, после его разделения, дает начало все же целостному образованию. Однако зачатки никогда не бывают вполне индифферентными. Они по меньшей мере обладают известной полярной структурой, некоторым «градиентом». Естественно, что и они допускают возможность различных сдвигов без нарушения нормального формообразования. И изменение интенсивности самого градиента может оказаться без заметного влияния. Таким образом, и «индифферентные» зачатки, выступающие в роли компонентов более сложных морфогенетических систем, обладают также способностью к регуляторному восстановлению своей структуры при ее нарушении.
В процессе эволюции развивается, таким образом, целая система регуляторных механизмов, «защищающих» нормальное течение процессов формообразования от всевозможных случайных нарушений. Все эти изменения идут, очевидно, под направляющим влиянием стабилизирующей формы отбора, основывающейся на постоянной элиминации особей, формообразование которых было существенно нарушено в результате случайных внешних влияний или в результате мутаций.
Д. ЗНАЧЕНИЕ РЕГУЛЯТОРНЫХ КОРРЕЛЯЦИЙ В ЭВОЛЮЦИИ
В предыдущем изложении я уже неоднократно отмечал значение регуляций как факторов, способствующих переживанию особей в разнообразных условиях существования, содействующих накоплению скрытого резерва наследственной изменчивости и поддерживающих пластичность организмов в процессе их исторического преобразования. Так как эти моменты имеют очень большое значение и последовательно вытекают из всего рассмотренного материала, то я нахожу нужным обобщить здесь все существенное, прежде чем перейти к основным выводам относительно эволюции индивидуального формообразования, которым посвящена следующая глава.
Вопрос о значении корреляций в эволюции был впервые поднят А. Н. Северцовым (1914), а затем разработан мною (1938а, 19386, 1939а,,19396, 19406). Здесь выдвигается главным образом вопрос об их значении на низовых этапах эволюции в самой общей форме. Более специальное освещение он получил в моей книге «Организм как целое в индивидуальном и историческом развитии».
а. Физиологическая эврибионтностъ. Установление регуляторных механизмов, противодействующих нарушениям формообразования со стороны случайных изменений в факторах внешней среды, означает ограничение способности к модификационным реакциям известными пределами одной или нескольких адаптивных норм.
Авторегуляция, в нашем определении, ведет к ограничению одной или нескольких «нормальных» реакций известными пределами интенсивности внешних факторов между нижним и верхним порогами реактивности тканей. В процессе эволюции оба порога реактивности раздвигаются шире, и если они вначале охватывают лишь область обычных небольших колебаний в интен-сивностях факторов внешней среды, то в дальнейшем они распространяются на более широкие пределы встречающихся иногда случайных уклонений. Для организма это означает повышение устойчивости формообразования и прогрессивную автономизацию развития. Об автономном развитии мы говорим в том случае, когда оба порога реактивности тканей раздвинуты за пределы встречающихся различий в факторах среды. Тогда нормальная реакция наступает при всех условиях и протекает исключительно под направляющим влиянием внутренних факторов развития (которые получают доминирующее значение и при авторегуляции).
Организм внешне не реагирует на изменения в факторах среды. Это объясняется, однако, исключительно существованием внутренних регуляторных механизмов, противодействующих внешним влияниям. Вместо морфогенетических реакций приспособительного характера при автономном развитии устанавливаются физиологические регуляции, нейтрализующие вредные влияния на развитие организма. Иными словами, морфологические адаптации заменяются физиологическими.
Физиологическая приспособляемость свойственна в известной степени всем организмам. Она возникала одновременно с самой жизнью и развивалась прогрессивно вместе с усложнением жизненных процессов. Установление авторегуляторных форм индивидуального развития, происходившее также уже на заре органической жизни, означает усиление физиологической приспособляемости организма. Собственно авторегуляция нескольких адаптивных норм достигает наивысшей ступени у высших растений. У животных авторегуляция ограничивается обычно одной основной адаптивной нормой, но дополняется весьма развитой способностью к функциональным адаптациям. Авторегуляция одной нормы достигает высших форм автономного развития только у животных. Очень совершенная система физиологических адаптации и регуляций животных усложняется еще развитием адаптивных форм движения, руководимых нервной системой (таксисами, инстинктами и, наконец, высшими, сознательными проявлениями деятельности нервной системы). Физиологическая и, в частности, нервная форма индивидуальной приспособляемости достигает у позвоночных, и особенно у высших позвоночных, наибольшего совершенства.
Физиологическая приспособляемость к изменениям во внешних факторах способствует широкому расселению и завоеванию новых мест в природе еще в гораздо большей степени, чем морфологическая приспособляемость, так как она устанавливается с гораздо большей скоростью и отличается быстрой и легкой обратимостью реакций.
Поэтому физиологическая приспособляемость связана с более широкой «эврибионтностью», чем морфологическая приспособляемость.
Организм может жить и размножаться в весьма разнообразных условиях. При быстроте физиологической приспособляемости, и в особенности приспособляемости поведения, организм относительно легко перестраивается даже при изменении (в известных пределах) биотической обстановки. При прочих равных условиях это означает уменьшение интенсивности элиминации и увеличение численности особей данного вида и более широкое его распространение (конечно, в пределах возможности физиологической адаптации данного вида).
Жизнь большой популяции в разнообразных условиях способствует также развитию генетической гетерогенности — накоплению резервов внутривидовой изменчивости, которая, благодаря наличию весьма совершенной системы морфогенетических регуляций, имеет в основном скрытые формы. Свободная миграция и скрещивание особей различных популяций способствуют возникновению все новых комбинаций. Тому же благоприятствует и малая интенсивность элиминации индивидуальных уклонений, компенсируемых способностью к физиологической адаптации. Все это означает наличие весьма благоприятных условий для накопления большого резерва наследственных изменений и для мобилизации этого резерва при изменении внешних условий. Поэтому высокое развитие регуляторных корреляций, всегда сопровождаемое физиологической приспособляемостью организма, оказывается важным фактором, повышающим эволюционную пластичность, т. е. мобильность вида в процессе его исторических преобразований. Потенциальная способность организмов к прогрессивной эволюции достигает в этом случае наивысшего уровня.
б. Прогрессивная интеграция органических форм. Если в процессе морфологической адаптации организма к разным сторонам внешней среды создаются новые дифференцировки, то через ре-гуляторные системы и физиологические адаптации эти дифференцировки связываются в гармонически построенное и согласованно действующее целое. Создается особая пластичность организации, при которой как благоприятные наследственные изменения, так и функциональные адаптации сразу сопровождаются согласованной с ними перестройкой всей организации. Адаптации дополняются уже в процессе индивидуального развития соответственной коадаптацией органов. Таким образом, организм эво-люирует всегда как гармоническое целое и отдельные новопри-обретения не нарушают этой согласованности организации. Новые функциональные дифференцировки фиксируются, однако, как наследственное усложнение организации, лишь через стабилизирующую форму естественного отбора. Только отбор наследственных уклонений, связанных с дальнейшей дифференциацией внутренних факторов развития (определяющих развитие этих дифференцировок), может привести к такому фиксированию. Это означает, однако, дальнейшее усложнение системы корреляций.
Таким образом, в процессе эволюции наравне с прогрессивной дифференциацией идет всегда и сопутствующая ей интеграция. Происходит объединение новых дифференцировок путем их подчинения общей системе внутренних факторов индивидуального развития. Это означает усложнение системы морфогенети-ческих взаимозависимостей, а вместе с тем и прогрессивную автономизацию онтогенеза (Шмальгаузен, 1938а, б, 1942).
5. ЭВОЛЮЦИЯ ИНДИВИДУАЛЬНОЙ ПРИСПОСОБЛЯЕМОСТИ И ФОРМООБРАЗОВАНИЯ
Если результаты физиогенной и в особенности функциональной адаптации приобретают постоянное значение в известных условиях существования, то они включаются в организацию и фиксируются в результате непрерывного действия стабилизирующего отбора. Вместе с тем происходит усложнение системы морфогенетических корреляций как внутренних факторов индивидуального развития, переходящего от зависимых дифференци-ровок к установлению все более автономных самодифференцирующихся систем.
Из этого вытекает значение индивидуального приспособления как фактора, намечающего дальнейший путь эволюции. Однако индивидуальная приспособляемость и сама меняет в процессе эволюции свои формы. Способность к физиогенным адаптациям дополняется способностью к функциональным и замещается затем, по мере развития регуляторных механизмов, способностью к физиологическим адаптациям. Среди последних у позвоночных животных выдвигается приспособительное поведение как высшее выражение регуляторных функций центральной нервной системы.
А. ИНДИВИДУАЛЬНАЯ ПРИСПОСОБЛЯЕМОСТЬ КАК ЭЛЕМЕНТ ОНТОГЕНЕЗА
Индивидуальное развитие всегда является частично зависящим от факторов внешней среды. Провести определенную грань между признаками и дифференцировками, зависящими и не зависящими от внешних факторов, практически невозможно — онтогенез в этом смысле неразложим. Однако несомненно, что удельный вес таких зависимостей у различных организмов весьма различен, и если мы не можем учесть долю участия различных факторов в нормальном онтогенезе, то можно установить влияние изменений внешних факторов на формообразование организма в условиях эксперимента. Таким образом, не только вскрывается различный масштаб этой зависимости, но и различное ее выражение у различных организмов. В огромном большинстве случаев, если внешние факторы меняются в тех пределах, в которых они изменяются в естественных условиях местообитания данного организма, зависимые изменения оказываются адаптивными. Так как в природных условиях каждая особь любого вида организмов подвергается тем или иным внешним воздействиям (т. е. их изменениям по сравнению со средней нормой), то это всегда оказывает известное влияние на индивидуальное формообразование. Кроме физиогенных модификаций при этом сказывается также и значение функциональных адаптации. Каждая особь оказывается в том или ином направлении модифицированной. Таким образом, адаптивные модификации входят в состав онтогенеза любой особи — они в большей или меньшей степени определяют конкретное ее оформление, являясь как бы индивидуальной надстройкой, известным завершением ее формообразования.
Масштаб и формы этой индивидуальной приспособляемости у различных организмов весьма различны и зависят от конкретных условий их существования, а также от формы и степени активности самих организмов.
У неподвижно прикрепленных организмов приспособляемость имеет почти исключительно физиогенный характер и выражается главным образом в изменениях роста отдельных частей, в частности — в явлениях неравномерного роста, ведущего к изгибам и определенной ориентировке по отношению к внешним факторам («тропизмы»). Это наблюдается у растений, а также у сидячих животных (губки, гидроиды и др.). У растений такие модификации выражаются и в изменении формы и строения органов (световая и теневая формы листьев, строение тканей листа, величина устьиц, толщина эпидермиса, развитие опушения, ветвление, развитие механических тканей стебля и мы. др.). Способность к физиогенным адаптациям достигает именно у растений своего высшего выражения, что находится в связи с пассивными формами распространения (ветром, животными), при которых семена при их прорастании совершенно случайно попадают в те или иные условия освещения, влажности, состава почвы и т. п. Сидячих животных это не касается в такой мере, так как у них всегда имеется стадия подвижной личинки, которая прикрепляется обыкновенно лишь на более подходящих местах.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 361; Нарушение авторских прав?; Мы поможем в написании вашей работы!