КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Возникновение приспособлений (адаптациогенез) 4 страница
|
|
|
|
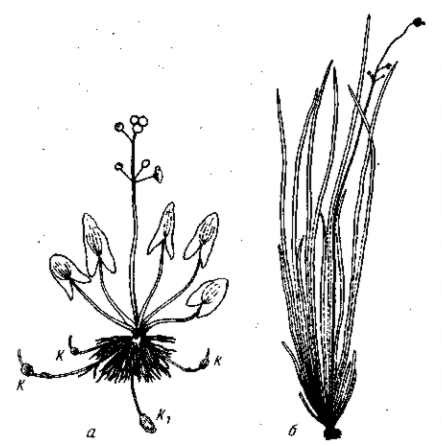
Рис. 32. Вполне развитые формы (адаптивные нормы) стрелолиста. По Schenck, 1915. а — наземная, б — водная форма
а. Модификационная эврибионтностъ. Способность к адаптивным модификациям означает всегда расширение среды обитания для данного вида организмов (или распространение на более значительный период времени в году — при сезонном полиморфизме). Организм может при соответствующих модификациях существовать и размножаться не в одном биотопе, а в двух или нескольких. Это означает но меньшей мере увеличение численности особей данного вида. Нередко это ведет и к более широкому распространению (в пределах возможной аккомодации). Вместе с тем это означает и уменьшение интенсивности элиминации, так как способность к индивидуальной адаптации предполагает и возможность приспособления к временным изменениям внешних факторов в пределах каждого из биотопов. Наконец, возможность известной адаптации и коадаптации означает до некоторой степени и сглаживание того нарушения соотношений, которое вносится теми или иными мутациями. Это связано опять-таки со снижением интенсивности элиминации.
Таким образом, в случаях развитой способности к адаптивным модификациям мы имеем (при прочих равных условиях) наличие более многочисленной популяции данного вида, живущей в более разнообразных условиях (биотопах), более сплошными массами (частично отпадает экологическая разграниченность биотопов) и в условиях менее острой элиминации физическими факторами ^при функциональных модификациях это касается и биотических факторов). Одним словом, результатом этой способности будет более или менее ярко выраженная эврибионтность, которую в этом случае можно ооозначить как «модификационную» эвриоионтность в отличие от «физиологической» эвриоионтности, о которой речь будет в дальнейшем.
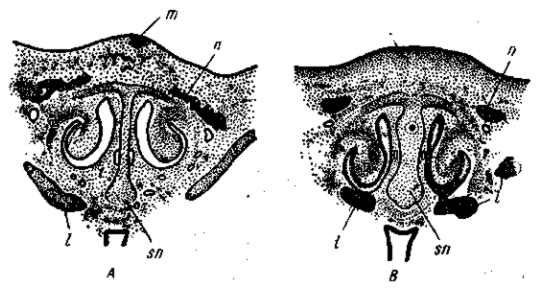
Рис. 33. Закладка обыкновенного гребня (А) у 8-дневного зародыша домашней курицы и розовидного (В) у 8,5-дневного зародыша породы виандот одинаковой стадии развития. Поперечные разрезы на уровне слезно-носового канала. Закладка розовидного гребня захватывает большую площадь покровов и значительную массу мезенхимы. Одновременно развивается более широкая носовая перегородка (sn). Закладка носовых костей запаздывает; с — обонятельный мешок, I — слезно-носовой канал, m — закладка гребня, п — закладка носовых костей
Значение морфологической приспособляемости организма не исчерпывается, однако, достижением более широких возможностей существования и размножения. Мы знаем, что в больших популяциях идет более легкое распространение и накопление мутаций и более свободное их комбинирование (панмиксия). Малая интенсивность прямой элиминации индивидуальных уклонений способствует также свободному накоплению многих мутаций и большему разнообразию возможных комбинаций.
Вразнообразных условиях существования поддерживается всегда и известный гетероморфизм, т. е. генетическое многообразие форм.
Таким образом, лабильность организации, т. е. способность организма к приспособительным модификациям, естественно связывается всегда и с эволюционной пластичностью, т. е. мобильностью вида в процессе исторического преобразования наследственных форм. В этой повышенной пластичности, т. е. потенциальной способности к прогрессивной эволюции, мы и видим наиболее общее выражение значения индивидуальной приспособляемости организмов.
6. Смена климатических зон и биотопов. Способность к адаптивным модификациям означает некоторую потенциальную способность к миграции за пределы первоначального местообитания. Климатические факторы не отличаются постоянством, и если организм может приспособляться к временным их изменениям, то он способен мигрировать в такие климатические зоны, в которых эти случайные уклонения в факторах среды приобретают постоянное значение. Так, например, организмы, живущие в местности, поражаемой иногда засухой, и приспособляющиеся к этим условиям, могут перейти в более засушливую область, где такие засухи представляют нормальное явление. Организмы резко континентального климата, приспособленные к жизни в весьма различной температуре (эвритермные), могут легко перейти к жизни в более холодном или более теплом поясе страны с менее континентальным климатом. Точно так же возможна в известных пределах и миграция из одного биотопа в другой. В особенности легко осуществляется, конечно, переход из общей среды в частную. Если организм индивидуально приспособляется к жизни в нескольких оиотопах, то он легко может осесть в любом из этих биотопов. Конечно, это означает специализацию, т. е. превращение эврибиоита в стенобионта. Такой процесс идет с большой легкостью.
При исторических сменах климатов, с их экологическими и биоценотическими последствиями, индивидуальная приспособляемость приобретает исключительное значение, организм, обладающий способностью модифицироваться соответственно локальным или сезонным изменениям в факторах внешней среды, оказывается вполне приспособленным, если подобные изменения становятся постоянной осооенностыо среды. Условно можно было бы говорить о преадаптации, если бы этот термин не связывался с представлениями известной теории Кено. Здесь по сути имеет место, конечно, не предварительное приспосооление, а всегда лишь приспосооление к реальным условиям существования данного времени. Однако условия, которые в данную эпоху или в данной области встречаются лишь локально, время от времени или периодически, могут в следующую эпоху распространиться на новые области или приобрести постоянное значение. Тогда прежняя второстепенная адаптивная норма может приобрести значение главной или даже единственной нормы. Такое «приспособление» организма происходит с огромной скоростью — в течение развития одного поколения. Вслед за этим произойдет, конечно, и наследственная перестройка организма соответственно его новому положению во внешней среде. Это осуществляется исподволь в процессе стабилизирующего отбора. В данном случае адаптивная модификация получает ведущее значение. Именно конкретная адаптивная модификация намечает путь дальнейшей эволюции. Во всех этих случаях «приспособления» не возникает, однако, ничего нового. Организм отвечает на изменения во внешней среде определенной реакцией, способность к которой уже давно приобретена в течение предшествующей эволюции данного организма в изменчивой внешней среде.
В этой связи мы приписываем особое значение функциональным модификациям, так как они отличаются большой гибкостью и, кроме того, определяются не столько изменением физических факторов внешней среды, сколько изменением биотической обстановки. Наконец, в этом случае приобретает особую роль активность самого организма. При изменении внешней обстановки, а также в процессе эволюции организма, он активно меняет свое положение во внешней среде. Млекопитающие, переходящие в поисках пищи из леса в степь, встречаются с иными врагами и не имеют иных средств защиты от больших хищников, кроме быстрого бега (крупные животные) или зарывания в землю (мелкие). Это означает для конечностей смену функций, а для всего огранизма — сложный комплекс функциональных модификаций. Млекопитающее, живущее в лесу, может находить защиту на деревьях. Это связано с приспособлением конечностей для лазанья. Широкая способность к функциональным модификациям во всяком случае значительно облегчает такой переход к древесному образу жизни. Следовательно, на базе функциональных модификаций могут возникать и совершенно новые приспособления.
Способность к функциональным модификациям оценивается нами очень высоко. Мы полагаем, что на базе общих реакционных способностей могут выявляться все новые частные реакции, дающие действительно новое, не только количественно, но и качественно новое оформление приспособляющемуся организму. Таким образом, адаптивная модификация может выходить и за пределы общей приспособленности реакций, приобретенной в предшествующий период эволюции. Она может вести к новым частным дифференцировкам, которые получают такую же ведущую роль в дальнейшей наследственной перестройке организма.
в. Возникновение новых функциональных дифференцировок. Главным процессом прогрессивной эволюции организмов является дифференциация, основанная на разделении функций и состоящая в расчленении однородного целого на разнородные части. Дифференциация органически связана и с противоположным процессом интеграции (см. ниже, гл. 4, а также И. Шмальгаузен, 1938а, 1942). Она означает прогрессивное «освоение» факторов внешней среды. Организм отвечает все более дифференцированной реакцией на внешние факторы и, с другой стороны, по мере «освоения»!, воспринимает все более тонкие градаций этих факторов, устанавливая между ними и качественные различия. Он дифференцируется, и по мере этой дифференцировки увеличивается качественное многообразие воспринимаемых факторов внешней среды. Взаимоотношения между организмом и средой все более усложняются. Определяющим моментом в прогрессивной эволюции организма являются устанавливающиеся конкретные взаимоотношения между организмом и средой, выражающиеся в борьбе за существование и ведущие через естественный отбор к преобразованию организма. В эволюции низших форм жизни ведущим процессом является, очевидно, освоение все новых сторон внешней среды и вместе с тем изменение и усложнение среды, определяемое в свою очередь размножением и увеличением многообразия органических форм, усложнением связей и взаимоотношений, в конкретных биоценозах. По мере прогрессивной эволюции организма увеличивается, однако, его активность, и ведущая роль в изменении взаимоотношений с внешней средой переходит к самому организму, все более дифференцирующему свои реакции, охватывающему новые факторы среды, активно мигрирующему в новые биотопы, активно вырабатывающему новые средства защиты и активно переходящему, по мере необходимости, на новое питание. В этом случае значение индивидуальной приспособляемости (способности к аккоммодации) непрерывно возрастает. В частности, возрастает значение функциональных модификаций и в особенности возрастает значение физиологических аккомодаций и приспособительного поведения.
В функциональных модификациях возникают и новые соотношения частей, создаются новые нормы. Переход к иному образу жизни (или на деревьях и т. д.), к иному способу передвижения (бег, лазанье, плаванье и т. д.), к иной пище (питание насекомыми, водными животными, растениями, семенами и т. д.), к иным способам добычи пищи (подстерегание, хватание на бету или на лету челюстями или лапами и т. д.), к иным способам защиты (бегом, зарыванием, вооруженной защитой или пассивной и т. д.) связан с различной активностью тех или других органов и ведет отчасти непосредственно к функциональной перестройке организма (Шмальгаузен, 1942а, 1939а). Экспериментальные данные показывают возможность такой перестройки (рис. 29). Что же касается масштаба перестройки, достигаемой путем аккомодации, то это вопрос совершенно второстепенный. Важно то, что эта перестройка достигается очень быстро — в течение одного онтогенеза, т. е. гораздо быстрее любого изменения внешней среды или проникновения вида в новый биотоп. Уже небольшое приспособительное изменение ставит организм в благоприятствуемое положение, и если стабилизация новых соотношений произойдет с достаточной скоростью (а стабилизирующая форма отбора идет, очевидно, с гораздо большей скоростью, чем ведущая форма естественного отбора), то на этой базе возможна и дальнейшая адаптивная модификация, как следующая ступень приспособления вида к новым условиям существования. На основе смены функций возникают, таким образом, новые модификации, которые являются ведущими в дальнейшей, более медленной наследственной перестройке организации.
Однако мы должны здесь подчеркнуть и возможность возникновения совершенно новых дифференцировок в результате индивидуальной аккомодации организма. Все новые дифференцировки связаны с разделением функций и определяются различным положением частей по отношению к внешней среде или к другим частям того же организма. Именно на этой почве происходили все прогрессивные изменения организации. Конечно, всегда остается законным вопрос: насколько такие именения определяются непосредственно модифицирующим влиянием самой функции, насколько — отбором новых, более дифференцированных форм реакций на эту функцию и насколько — отбором «независимых» наследственных изменений данной структуры. Несомненно, это бывает различно в отдельных случаях. Мы отмечаем, однако, принципиальную возможность непосредственного влияния функции и возрастающее значение этого влияния по мере установления высших, более активных форм жизни, характеризующих эволюцию наземных позвоночных.
Активность высших животных определяется в значительной мере дифференциацией нервной и мышечной систем. Все остальное имеет подчиненное значение. Однако как раз в отношении нервной и мышечной систем роль тренировки исключительно высока. У птиц и, особенно, у млекопитающих тренировка нередко входит как обязательный элемент онтогенетического развития молодого животного (через игры и «обучение»). Мы знаем, что дифференциация функций между частями одной мышцы приводит всегда к их обособлению и, во всяком случае, комплекс однородных мышечных пучков может распасться на разнородные, вполне самостоятельные мышцы (такова филогенетическая история осевой мускулатуры позвоночных, мускулатуры конечностей).
История органов чувств показывает их постепенную дифференциацию из общих покровов, а вместе с тем и обособление соответствующих нервных центров. С другой стороны, и дифферен-цировка мышечной системы сопровождается усложнением строения нервной системы. Наконец, и многие преобразования в скелете зависят от функции связанных с ними мышц. Частично все эти изменения строятся за счет функциональных модификаций. Это доказывается экспериментами выключения нормальной функции мышц или нервов, ведущими к частичному недоразвитию скелетных и мышечных образований. Многие такие изменения наблюдаются уже при содержании диких зверей в неволе. Если способность к функциональной гипертрофии в наших экспериментах и оказывается ограниченной, то вое же она настолько велика, что может обусловить неизмеримо более значительные преобразования в течение жизни одного поколения, чем самый интенсивный отбор наследственных изменений на протяжении существования многих сотен поколений.
При функциональной адаптации, связанной с установлением новых дифференцировок, эти изменения строятся, конечно, на основе наследственной, т. е. выработавшейся в предшествующей истории организма, нормы его реакций. Способность мышц к усилению в результате тренировки, способность нервных центров к некоторой гиперплазии, способность скелетных частей усиливаться и перестраиваться соответственно измененной нагрузке и все другие выражения функциональной гипертрофии основываются на унаследованной норме реакций данного организма. Она определяет и диапазон возможных изменений и, следовательно, границы возможной индивидуальной приспособляемости. Однако унаследованной оказывается лишь данная реакционная способность в общем — для мускульной ткани, для нервной ткани и для скелетных образований. Вполне, конечно, возможны различия в реакционной способности тех или иных категорий мышц (мышц конечностей, осевых или висцеральных мышц), областей нервной системы (первичных двигательных ядер и высших областей первичных и вторичных чувствующих центров) и разных скелетных структур (пластинчатые, трубчатые и компактные кости). Однако совершенно невероятным было бы предположение, что каждая отдельная мышца обладает своей строго определенной способностью к тренировке. Многие различия определяются, очевидно, только тренировкой, только реальным распределением функций в организме, находящемся в данной конкретной обстановке.
В некоторых случаях удается установить значение функции даже в развитии «пассивных» образований. Известно, что кожа млекопитающих реагирует на постоянное давление или трение образованием мозолей. Это — общая реакция, которая, однако, приобретает иногда значение постоянной местной реакции (например, подошвенные мозоли на лапах многих млекопитающих). И у человека на подошвах ног кожа значительно утолщена. Свинья-бородавочник (Phacochoerus) разрывает землю для добычи корма и при этом всегда длительно опирается на локтевой сустав. Соответственно на локтях у нее развиваются постоянные мозоли. Так как эти мозоли, очевидно, появились в результате новой привычки (а не наоборот,— нельзя допустить, что свинья стала рыть землю в такой позе именно потому, что у нее развились мозоли), то первоначально они имели характер чисто модификационных изменений, возникших на базе общей реакционной способности кожи. Здесь произошла, следовательно, явная дифференциация — на базе исторически развившейся общей реакционной способности выделилась специальная реакция местного характера (местная мозоль) как приспособление к особым условиям существования Именно это частное модификационное изменение приобрело ведущее значение в дальнейшей эволюции. В процессе эволюции оно стабилизировалось как наследственное изменение. Исследования Лехе (Leche, 1902) показали, что эти мозоли закладываются уже у зародыша. Точно так же указывают, что и у человеческого зародыша эпидермис на подошве ног оказывается уже утолщенным. Следовательно, то образование мозолей, которое возникло первоначально как модификационное изменение в виде ответа ' на местное раздражение, стало затем развиваться на соответствующем месте и без этого внешнего раздражителя, под влиянием внутренних факторов (автономизация развития). Это означает стабилизацию данного новообразования, возникшего под непосредственным дифференцирующим влиянием функционального раздражения.
г. Стабилизация конкретных модификаций. Если при переходе из одного биотопа в другой физиогенная модификация имеет скачкообразный характер замены одной хорошо выраженной адаптивной нормы совершенно иной (водная форма, ксерофитная, альпийская и т. п.), то это, конечно, не составляет общего правила. Гораздо чаще историческое изменение условий существования совершается более постепенно и сопровождается таким же постепенным приспособлением организма.
Особое значение имеют при этом общефизиологические и функциональные модификации. Они реализуются в пределах существующих общих норм реакций в виде дифференцированных частных реакций.
Стабилизация конкретных результатов таких реакций должна происходить во всех тех случаях, когда данная модификация становится общим достоянием целой популяции в ряде поколений. Мы знаем, что стабилизирующая форма отбора осуществляется на базе селекционного преимущества приспособленной (в том числе и вновь приспособленной) нормы перед всеми от нее уклонениями, т. е. путем накопления всех мутаций, не выводящих фенотип за пределы установившейся нормы.
Можно, конечно, предполагать и одновременное действие обычной формы естественного отбора в том же направлении, в каком происходит модифицирование организма (на основе селекционного преимущества дальнейших отклонений по сравнению с уже осуществленной нормой). Это возможно, если модификация недостаточна, мало устойчива (в данных условиях) или реализуется лишь поздно в онтогенезе данной формы. На такую замену модификаций наследственными изменениями, обладающими известными преимуществами, обратил внимание в особенности Лукин (1935, 1936, 1940). Я не отрицаю этих процессов. Однако это не есть особая форма естественного отбора. Поскольку отбор основывается в этом случае на положительных наследственных изменениях, а возможность таких изменений довольно ограничена, то это, очевидно, весьма медленный процесс (ведущая форма отоора). С другой стороны, стабилизирующий отбор не идет далее установившейся нормы и основывается не на положительных уклонениях, а на «нейтральных» изменениях (т. е. не только на безразличных, но и на частично и условно вредных мутациях, вредные выражения которых нейтрализуются; главным же образом вообще на малых мутациях). Эти мутации встречаются неизмеримо чаще и, главное, свободно размножаются, накапливаются и комбинируются в популяциях, в пределах нормального «дикого» типа. В мобилизационном резерве внутривидовой изменчивости имеется всегда готовый запас таких мутаций и комбинаций, которые, оставаясь в пределах нормы, оказываются в данных условиях более жизнеспособными, т. е. оставляют более значительное потомство, чем любые отклонения от нормы (независимо от того, каковы источники этих уклонений). Только это и составляет основу процесса стабилизации. Особи с наследственным уклонением, реагирующие на случайные факторы внешней среды возвратом к прежней норме, утратившей свое значение, элиминируются. Этим путем размножаются более устойчивые комбинации, «фиксирующие» достигнутый уровень приспособления. На основе этой новой нормы возможны дальнейшие конкретные модификации, которые по мере приобретения ими постоянного значения (в переживании особей данной популяции) в свою очередь стабилизируются в виде дальнейшей нормы, которая может служить вновь исходной позицией для прогрессирующего приспособления. Таким образом, я вовсе не думаю, что сначала происходит модификация в полной мере ее потенциальной возможности, затем происходит ее дальнейшее совершенствование через обычный отбор и, наконец, фиксирование через стабилизирующий отбор.
Я представляю себе непрерывную кооперацию этих процессов. С одной стороны, организм непрерывно до известной степени модифицируется, и, с другой стороны, те выражения индивидуальной и, в частности, функциональной адаптации, которые уже приобрели постоянное значение в данных условиях, непрерывно стабилизируются (через элиминацию фактических уклонений). Это означает сужение нормы реакций в известном направлении, утратившем свое значение. Однако через ведущую форму отбора норма реакций меняется в иных направлениях. Устанавливаются, быть может, некоторые новые адаптации, а вместе с тем расширяется в определенном направлении, диктуемом реально встречающимися факторами внешней среды, и способность к дальнейшим адаптивным модификациям (расширение нормы реакций и лабилизация формообразования).
Континентальный климат, с господством резких колебаний влажности, инсоляции и, в особенности, температуры, должен способствовать выработке наиболее стабильных форм, обладающих наиболее высоко развитой системой регуляторных механизмов. Поэтому мы считаем весьма вероятными известные гипотезы, связывающие эволюцию покрытосеменных растений, с одной стороны, и млекопитающих животных, с другой, с засушливыми периодами мезозоя и с континентальными климатическими зонами земной поверхности.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 493; Нарушение авторских прав?; Мы поможем в написании вашей работы!