КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
ВІДПОВІДІ 1 страница
|
|
|
|
КОМПЛЕКСНА КОНТРОЛЬНА РОБОТА
КОМПЛЕКСНА КОНТРОЛЬНА РОБОТА
з дисципліни «Основи педагогіки та методика викладання»
(назва дисципліни)
до акредитації напряму
8.03060101 «Менеджмент»
Студент _________________________________________________________________
(прізвище, ім’я, по батькові)
Курс ______________________________
Група ______________________________
Початок роботи _______ год. _______хв.
Закінчення роботи ______ год. ______хв.
Питання контрольного завдання№24
1. Цілі, завдання, принципи виховання у вищій школі.
2. Функції процесу навчання, їх взаємозв'язок.
3. Задача: Одного разу Ви стали учасником дискусії викладачів з приводу того, якою мовою – українською або російською треба викладати у вищому навчальному закладі.
Запитання і завдання:
1) Яку точку зору викладачів Ви би підтримали?
2) Обґрунтуйте власну точку зору, наведіть переконливі документи.
| Оцінка | Викладачі | Експерти | ||
| Прізвище | Підпис | Прізвище | Підпис | |
з дисципліни «Основи педагогіки та методика викладання»
(назва дисципліни)
до акредитації напряму
8.03060101 «Менеджмент»
Студент _________________________________________________________________
(прізвище, ім’я, по батькові)
Курс ______________________________
Група ______________________________
Початок роботи _______ год. _______хв.
Закінчення роботи ______ год. ______хв.
Питання контрольного завдання№25
1. Засоби навчання. Традиційні засоби навчання. Місце і роль нових інформаційних технологій серед засобів навчання.
2. Методика читання лекцій у ВНЗ.
3. Задача: Між двома студентами виник конфлікт, який негативно впливає на позитивний психологічний клімат в академічній групі. Кожен із студентів звертається до викладача окремо по допомогу і підтримку його в конфліктній ситуації.
Запитання і завдання:
1)Проаналізуйте педагогічну ситуацію, сформулюйте основні суперечності, що виникли в ній.
2)Від чого бути залежати вибір стратегії Вашої поведінки в пошуках способів примирення конфліктуючих?
| Оцінка | Викладачі | Експерти | ||
| Прізвище | Підпис | Прізвище | Підпис | |
1.1.
Праці Гіппократа, Аристотеля й інших древньогрецьких філософів містять відомості екологічного характеру. Ще на початку 18 століття Антон Ван Левенгук був піонером у вивченні трофічних ланцюгів і регуляції чисельності організмів, а праці англійського ботаніка Ричарда Бредлі свідчать, що їх автор мав чітке уявлення про біологічну продуктивність.
Як самостійна наука екологія виникла на початку 20-го століття, проте її назва “екологія” увійшла до загального лексикону лише в останні десятиліття.
Рух, який можна назвати “загальна стурбованість проблемами довкілля” зненацька розгорнувся з 1968 по 1970 рр. Усі раптом зацікавилися забрудненням середовища.
До 1970 року екологія сприймалася, головним чином, як один із підрозділів біології. З цього часу екологія часто трактується як інтегрована, міждисциплінарна наука.
Останніми роками говорять вже про екологію слова, екологію думки, екологію будь-чого. Таким чином межі науки виявилися розмитими, а предмет розширився до практично повної невизначеності.
Якщо головним предметом екології визнати екосистему, а відтак екологічним вважати будь-яке дослідження, що має з’ясувати роль того чи іншого об’єкта або явища в екосистемі, то відразу припиняються безплідні дискусії про місце екології у системі наук. В усіх інших випадках межі екології лишаються невизначеними, а сама наука сприймається як щось комплексне, міждисциплінарне, тобто може тлумачитися будь-як.
У зв’язку з визначенням головного об’єкту екології зникає і проблема належності її до біологічних наук. Біологія вивчає живу форму руху матерії. Конкретні біологічні дисципліни значну увагу приділяють вивченню взаємозв’язків організмів із навколишнім середовищем. Але в усіх випадках абіотичні компоненти вони розглядають як зовнішні чинники щодо конкретного організму, популяції чи угруповання. Тобто предметом біологічних дисциплін залишається лише жива форма руху матерії. В екології, на відміну від біології, неживі компоненти екосистеми розглядаються як складові системи, тобто вони входять «повноправними» елементами до головного об’єкта екології. Саме тут проходить межа між біологічними й екологічними науками. Саме нерозуміння цього нюансу і породжує безліч дискусій як про предмет екології, так і про її місце в системі наук. А водорозділ між екологією і біологією досить тендітний: як тільки абіотичні компоненти ми включаємо до складу об’єкту, тобто переносимо з розряду “зовнішнє середовище” до розряду “склад системи”, тобто вони стають “повноправними” елементами системи, так відразу екологія виокремлюється з біологічних наук в самостійну.
Таким чином, екологія вивчає структурно-функціональну організацію різноманітних екосистем та процеси, що в них відбуваються з метою прогнозування подальших змін та оптимізації взаємин людини з Природою, що можливо лише на терені гармонізації всіх форм діяльності людини, і є надійною запорукою стабільного розвитку суспільства. І лише за умови усвідомлення себе частиною цього дивовижного розмаїття, а не на засадах постійного протипоставляння людини і природи, що призводить до безглуздих і водночас украй небезпечних спроб “підкорення” природи, наслідком чого є глобальне спотворення колись чарівних осередків життя, а зараз спустошених та занедбаних територій і акваторій, можливе виживання людини як виду.
І ще варто нагадати, що екологію не слід плутати з інвайроментологією (від англійського invironment) – наукою про середовище. Це дійсно, на даному етапі розвитку природничих наук, комплексна наука, що включає всі дисципліни, які торкаються окремих проблем довкілля. Екологія також вирішує багато проблем інвайроментології. Але сюди належать і такі науки, як гідрологія, гідрохімія, геоморфологія, кліматологія, ландшафтознавство тощо.
Визнання екосистеми центральним об’єктом екології вимагає застосування системного підходу і загальної теорії систем як головної методологічної основи вивчення екологічних процесів і явищ.
Часто екологію розуміють як науку про охорону довкілля чи природи. Це також не відповідає дійсності. Звичайно, для розробки заходів щодо охорони певних видів необхідно мати достатньо інформації про їхнє місце і роль у конкретній екосистемі, визначити ємність середовища та лімітуючі їхню чисельність фактори тощо.
Для вжиття заходів щодо зниження рівня забруднення середовища необхідна інформація, які саме забруднення викликають найістотніші негативні зміни в екосистемі, які шляхи їх циркуляції в екосистемі, де і в якій кількості вони накопичуються тощо. З іншого боку необхідна інформація про максимально припустимий рівень даного токсиканта (чи іншого забруднення) в екосистемі, який не викликає ніяких достовірних змін у структурно-функціональній організації екосистем і не має токсичного ефекту (з урахуванням дії інших факторів, явищ синергізму, кумуляції тощо).
Таким чином екологія є науковою основою всієї природоохоронної роботи, а також невід’ємною складовою екологічного менеджменту, екологічного прогнозування, оптимізації тих чи інших форм господарської чи іншої діяльності людини.
Щоб уникнути багатьох безплідних дискусійних питань (з проблем, які давно вже вирішені загальною теорією систем і ретельно розроблені в рамках системного підходу) та зрозуміти конкретні екологічні закономірності через призму загального прояву притаманних будь-яким системам властивостей, спочатку варто розглянути основні поняття системного підходу.
1.2.
Одним з центральних понять екології є поняття “екологічної ніші”. Воно є найбільш повною характеристикою місця і ролі конкретної біосистеми (популяції, геміпопуляції тощо) в екосистемі.
Вперше термін «ніша» в екологічній літературі використав Р. Джонсон (Johnson, 1910) у зв’язку з характеристикою залежності поширення видів від
Однак широке використання терміну «ніша» в екологічній літературі почалося після виходу у світ оригінальної роботи Джозефа Грінелла, присвяченій опису ніші, що зайнята каліфорнійським (пересмешником) (Toxostoma redivivum) (Joseph Grinell, 1917) в асоціації чапараллю. За Грінеллом, ніша пересмешника характеризується комплексом умов (абіотичні фактори, їжа, укриття тощо), які притаманні густим заростям кущів чапаралля, де тільки і можливе існування даного виду. І хоч сам Грінел чітко не сформулював визначення екологічної ніші, його розуміння ЕН охоплювало як абіотичні умови, так і біотичні фактори існування виду.
Подальший розвиток поняття ЕН знайшла у працях Чарльза Елтона “Екологія тварин” (Ch. Elton, 1927), та інших його публікаціях. Елтон одним з перших надав ніші сенсу “фунціонального статусу виду в угрупованні”, розвиваючи уявлення Ч.Дарвіна про спосіб життя виду, ролі, “професії” і його місці в економії природи. Так, за Елтоном, під нішею тварини розуміють спосіб життя і зокрема спосіб харчування в тому ж розумінні, в якому говорять про професію в людському суспільстві. При цьому підкреслюється, що ідея ніші має суто екологічний, а не таксономічний характер.
Якісно новий етап у розвитку поняття ЕН починається з 1957 р., коли Хатчінсон (G.E.Hutchinson,1957) вперше сформулював це поняття як суму усіх зв’язків організмів даного виду з абіотичними умовами середовища і з іншими видами живих організмів і показав, що нішу можна розглядати як багатомірний простір, або гіперпростір, в межах якого умови середовища допускають необмежено тривале існування особини чи виду.
Хатчінсон розрізняв фундаментальну нішу – гіперпростір, в межах якого можливе існування виду, коли він не обмежений конкуренцією з іншими - і реалізовану нішу – менший гіперпростір, який займає вид при біотичних обмеженнях.
Таке розуміння екологічної ніші наближається до поняття “елемент системи”. Іншими словами, вивчаючи будь-який біоценоз чи угруповання ми можемо розглядати екологічні ніші окремих видів як елементи угруповання. При цьому топічна ніша окреслює місце його в системі, а функційна ніша вже входитиме до структури угруповання.
Тоді весь біоценоз можна уявити як сукупність взаємопов’язаних між собою екологічних ніш. Адже кожний елемент системи має своє “місце” в системі і характеризується певним колом зв’язків як з іншими елементами системи, так і з зовнішнім (для даної системи) середовищем. У цьому випадку елементом системи (у більшості випадків) можна визнати саме геміпопуляцію.
Згідно принципу ієрархії рівнів організації внутрішня структура елемента системи не вивчається і не розглядається. Розуміючи багатомірну екологічну нішу як повну характеристику місця і функціональної ролі даної біосистеми в угрупованні чи екосистемі, треба мати на увазі кілька обставин. Перш за все необхідно з’ясувати, для біосистем якого рівня зазвичай слід використовувати саме поняття екологічної ніші. Найчастіше воно використовується для популяції. Саме поняття популяції в екології найчастіше використовується в розумінні самовідтворної одновидової системи в рамках даної екосистеми. Проте в безлічі випадків ми не можемо говорити, що популяція займає певну екологічну нішу. Личинки хірономід, куліцид, бабок, як і багатьох інших комах, є невід’ємними складовими (тобто елементами) різноманітних водних екосистем. Але імаго притаманне наземне середовище. Личинки безхвостих амфібій також займають зовсім іншу екологічну нішу, ніж дорослі форми. Так, ропухи мешкають на суходолі, в той час як їхні личинки ведуть виключно водяний спосіб життя. За спектром живлення дорослі ропухи - хижаки, що живляться наземними комахами, червами і навіть хребетними тваринами – пташенятами тощо. Личинки ж за характером живлення не мають з дорослими нічого спільного. Медузи і поліпи також істотно відрізняються за умовами свого існування і входять до складу різних біоценозів – планктону (медузи) та бентосу чи перифітону (поліпи). Досить істотно відрізняються ніші спорофіту та гаметофіту рослин. І таких прикладів можна навести безліч. Таким чином, у багатьох випадках для характеристики структурованості екосистем чи біоценозів за нішевим простором доцільно (а у багатьох випадках – необхідно) використовувати поняття геміпопуляції як розмірно-вікової групи певного виду, або конкретного етапу розвитку, що займає певну екологічну нішу. Таким чином, одна популяція може займати кілька екологічних ніш, значно розширюючи свою представленість в різноманітних екосистемах, що, безперечно, сприяє лише процвітанню даної популяції та й виду в цілому.
Таким чином, розуміючи екологічну нішу як місце і роль даного об’єкта в системі, ми можемо застосовувати це поняття для біосистем різного рівня організації – геміпопуляцій, популяцій, гільдій тощо.
Варто розрізняти фундаментальну (потенційну) та реалізовану ЕН. ПЕН окреслює можливий спектр мешкання певної спільноти, в той час як РЕН є характеристикою реалій стану біосистеми в даній екосистемі.
Багатомірна (n – мірна) екологічна ніша складається з n окремих ніш. Так, говорять про трофічну нішу, топічну нішу (яка в свою чергу значною мірою пов’язана з факторіальними нішами - термальною тощо) та інші. Так, при створені концентуальної моделі трофічної структури даної екосистеми її елементами будуть популяції, геміпопуляції або гільдії (в залежності від мети конкретного дослідження). Структура цієї системи буде визначатися особливостями трофічних зв’язків між її елементами.
Графічним зображенням трофічної структури найчастіше виступає зображення трофічної мережі у такому вигляді:
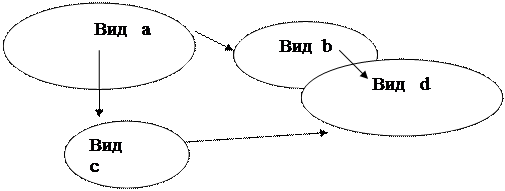
Звідки видно, які види поїдають яких, тобто яким шляхом відбувається трансформація речовини ї енергії в угрупованні.
А для характеристики ступеня схожості раціонів (трофічних ніш) найчастіше зображують таку схему:
А для перевірки досконалості цієї моделі доцільно застосувати балансовий підхід: інтенсивність вхідних і вихідних потоків між елементами системи мусить бути збалансованою. Так, у випадку стаціонарного стану системи, коли біомаса окремих її елементів відносно постійна, енергетичні потоки на вході до кожного елемента повинні бути близькими до енерговитрат цього елемента. У випадку дизбалансу, приріст (чи зменшення) біомаси чи енергії кожного елемента повинен дорівнювати різниці між вхідними та вихідними енергопортоками. У випадку виявлення значної невідповідності між ними необхідно провести додаткові дослідження по цим елементам. Причому схожість живлення тут представлена між усіма видами (дано перекриття раціонв кожного з кожним). Так, максимальна ступінь схожості раціонав між видами с і е (74 %), мінімальна у видів а і d (5 %). Але загальний внесок до дифузної конкуренції на трофічній ниві – це і буде сума всіх впливів видів – конкурентів (за живленням), але і це ще не вичерпує дифузної конкуренції. Конкуренції може бути і без перекриття трофічних ніш (скажімо, топіна – використання одних і тих же ділянок). Таким чином, навіть початок знайомства з дифузною конкуренцією показує, з одного боку, складність проблеми, але з іншого – цілком реальні шляхи до кількісної оцінки її загального впливу (за станом об’єкту, що досліджується).
Дві останні схеми також можуть бути ілюстрацією до поняття елемента угруповання. Часто постає питання: що є елементом трофічної структури біоценозу? Вочевидь, не популяція. Так, личинки хірономід і дорослі форми, пуголовки і жаби і таких прикладів маса. Аналогічно щодо топічної структури. Розглянемо, які біотопи займають різні вікові стадії вже згаданих істот. І у цьому випадку “популяцією” оперувати досить незручно. Як і у попередньому прикладі, без поняття “геміпопуляція” тут не обійтися. Врешті решт, аналізуючи схожі блоки проблем дійдемо висновку, що для характеристики структурованості угруповань і екосистем одним з найзручніших понять є “геміпопуляція”. Саме вона задовольняє вимогам до елемента системи і, з іншого боку, згідно концепції рівнів ієрірхії системи елемент її розглядається за принципом “чорної скриньки”.
Таким чином, послідовно застосовуючи принципи системного підходу до вивчення екологічної ніші, можна розглядати її саме як місце даного елемента в екосистемі, а сукупність усіх зв’язків елемента з іншими і з зовнішнім середовищем як роль елемента в системі. На практиці частіше може застосовуватися схема певного типу структури (трофічної, топічної, енергетичної, інформаційної, біогеохімічної тощо), де кожний елемент (зокрема, геміпопуляція) має своє місце і структуру внутрішньо- і зовнішньосистемних зв’язків).
Найчастіше він формулюється так: види, що займають одну й ту ж екологічну нішу, не можуть стійко співіснувати. Один вид неминуче витіснить інший.
Класичною працею, що підтверджує цей принцип, вважається робота Г.Ф.Гаузе (1934), де в результаті експериментального вивчення конкуренції двох видів інфузорій – Paramecium caudatum і P. aurelia було показано, що один вид витісняє інший. Інфузорій цих видів культивували роздільно і разом, використовуючи в якості їжі бактерій Bacillus pyocyaneus, яких давали щоденно в певній кількості. За умов роздільного культивування обидва види демонстрували типовий S-подібний ріст (мал.) При сумісному утриманні кожний вид спочатку швидко збільшував свою чисельність, сумарна біомаса обох видів досягала максимуму, а потім вже починалося власне конкурентне витіснення, зазвичай це закінчувалося перемогою P. aurelia.
Необхідно зазначити, що процес конкурентного витіснення в даному випадку цілком визначався різною швидкістю розмноження інфузорій, що компенсувало регулярне вилучення (експериментатором!) певної кількості особин обох видів. Таке вилучення в обсязі 0,1 від загального об’єму води (а відповідно, і кількості інфузорій кожного виду) проводилось щоденно з метою підтримання постійного росту популяцій і уникнення досягнення стаціонарного стану, який, як вважав Г.Ф.Гаузе, міг би істотно вплинути на важливі особливості популяцій.
Поразка P. audatum в конкуренції пояснювалалася тим, що вона погано переносила накопичення в середовищі шкідливих продуктів метаболізму бактерій. Якщо ж в якості їжі використовували дріжджі Sacchаromyces exiguus або бактерій іншої лінії B.pyocyaneus, а також якщо частіше промивали середовище, то перемагала в конкуренції P.caudatum, швидкість розмноження якої за даних умов була вищою. Закон Гаузе спричинив значний вплив на формування екологічної думки в 40-50-х роках.. Популярності його сприяло і те, що він відповідав правилу просторового розмежування близькоспоріднених видів, яке трактувалося натуралістами XIX-початку XX століття як засіб ослаблення міжвидової конкуренції. Зокрема американський натураліст Дж. Грінелл писав у 1904 році “лише пристосування до різної їжі чи способів її добування досягається те, що більш ніж один вид може займати одне місце мешкання. Малоймовірно, щоб два види приблизно однакових харчових потреб довго мешкали в одному районі. Один витіснить іншого (Grinell, 1904).
- Термін “екологія” (від грецького “ойкос” – будинок, житло, помешкання) запропонував у 1866 р. відомий німецький дослідник Ернст Геккель у праці “Загальна морфологія організмів”. Екологією він називав “загальну науку про стосунки організмів з навколишнім середовищем”. Згідно запропонованої Геккелем ієрархічної класифікації біологічних наук, екологія входила до складу фізіології і навіть іменувалася “фізіологією взаємовідносин”.
- Слід відзначити, що в екології однофакторні експерименти виявляються значно менш продуктивними, ніж у фізиці чи інших природничих науках. Адже коли проводиться однофакторний експеримент, скажімо, досліджується вплив температури на рівень фотосинтезу, то треба мати на увазі, що отримані в ньому результати справедливі лише для даного рівня освітлення, спектрального складу світла, концентрації окремих біогенних елементів, вологості тощо.
- За допомогою холістичного підходу одержують загальні параметри екосистемних процесів (що практично неможливо на засадах мерологічного підходу), а вже з’ясування механізмів окремих процесів досягається на терені мерологічного підходу. Так, вивчивши складові енергетичного балансу будь-якої екосистеми, корисно з’ясувати, внесок яких популяцій найбільш істотний у продукційно-деструкційні процеси.
- Патієнти – це види, здатні виживати в несприятливих умовах, де більшість інших видів існувати просто не здатні, наприклад за умов недостатнього освітлення, вологи, мінеральних речовин тощо. До патієнтів належить багато рослин, які вважаються посухостійкими. Тінелюбними чи навіть солелюбними. Хоч експериментально показано, що багато з них (хоч і не всі) за відсутності конкурентів можуть існувати і добре почуватися в умовах більшої вологості, освітленості тощо.
- Основний предмет екологічних досліджень – взаємозв’язки між живими організмами, їх групами різних рангів, живою і неживою компонентами екосистем, а також особливості впливу природних і антропогенних чинників на функціонування екосистем та біосфери в цілому.
2.1.
Системою можна назвати будь-який об’єкт, цілісні властивості якого є результатом взаємодії його складових.
У цьому визначенні у неявній формі є платонівське положення, що ціле є чимось більшим, ніж сума його частин. Причому якісно нові особливості цілого у порівнянні з його складовими є результатом взаємодії елементів системи між собою. Водночас система є якісно новим, своєрідним утвором у порівнянні з її елементами. В цьому знаходить свій прояв принцип емерджентності.
При вивченні ієрархічної організації систем у міру об’єднання компонентів, чи підмножин, у більш крупні одиниці, у них з’являються нові властивості, які не відмічалися на попередньому рівні організації. Такі якісно нові, емерджентні, властивості неможливо передбачити, виходячи з властивостей компонентів, що складають цей рівень організації. Так, властивості молекул безпосередньо не випливають з властивостей атомів, що входять до їх складу. Стосовно екологічних систем, то їхні властивості також неможливо передбачити, виходячи з нижчих рівнів організації.
Фейблмен (Feibleman, 1954) вважає, що при кожному об’єднанні підмножин у нову множину виникає не менше однієї нової властивості.
Солт (Salt, 1979) пропонує розрізняти емерджентні й сукупні властивості, що є сумою властивостей компонентів. І ті й інші – властивості цілого, проте сукупні властивості не мають нових чи унікальних особливостей. Так, біомаса популяції дорівнює сумі мас усіх особин, що входять до її складу
Емерджентні властивості саме й обумовлені взаємодією компонентів при формуванні системи певного рівня організації. На рівні популяції емерджентними будуть: її структура (статева, генетична, розмірно-вікова тощо), тип розподілу організмів у просторі, народжуваність, смертність, біотичний потенціал, тип динаміки тощо. На рівні угруповання – типи міжпопуляційних взаємин, трофічні ланцюги й мережі, видове розмаїття тощо. На екосистемному – характер біогеохімічних колообігів, трансформація енергії, енергетичний баланс екосистеми, екологічні сукцесії, флуктуації й трансформації тощо.
2.2.
Екосистемою можна назвати будь-який об’єкт, де живі і неживі компоненти функціюють як єдине ціле завдяки колообігу речовин, що здійснюється з використанням зовнішнього джерела енергії і призводить до створення певної структури, або, іншими словами – це система, елементами якої є біотичні і абіотичні компоненти, пов’язані речовинно-енергетичними та інформаційними потоками, відмежована від аналогічних утворів колообігом речовин певного ступеня замкненості.
Форбс (S. Forbs, 1887) першим розглядав природний комплекс організмів та їхнього абіотичного оточення як цілісну систему і запропонував для нього термін “мікрокосм”. Мікрокосмом він назвав озеро разом з організмами, що його населяють. Форбс підкреслював, що вплив на окремі компоненти неминуче тягне за собою і зміни всього цілого; водночас аналіз цілого є необхідною умовою задовільного розуміння будь-якої його частини. Таким чином “мікрокосм” Форбса досить близький сучасному поняттю “екосистема”.
В умовах водойм цілісна взаємодія біотичних і абіотичних факторів виявляється особливо чітко. Тому не випадково саме в гідробіології в кінці 19 – початку 20 століття почали успішно розроблятися проблеми взаємодії біоценозу і біотопу як компонентів єдиного цілого (Hensen, 1887; Джонсон, 1919).
Слід відзначити, що необхідність синтетичного підходу у вивченні наземних екосистем була вперше усвідомлена і сформульована у вигляді головної концепції вченими, які працювали над вивченням грунтів – природного тіла, в утворенні якого в нерозривній єдності сплелися біотичні і абіотичні фактори. Це відбулося в самому кінці 19 століття, коли з закликом розгорнути міждисциплінарні комплексні дослідження цілісних природніх систем виступив видатний російський вчений В.В.Докучаєв.
“Вивчалися, головним чином, окремі тіла, – мінерали, гірські породи, рослини і тварини, – і явища, окремі стихії, вогонь (вулканізм), вода, земля, повітря, в чому, повторюємо, наука і досягла, можна сказати, дивних результатів, але не їх відносини, не та генетичний, віковічний, і завжди закономірний зв’язок, що існує між мертвою і живою природою, між рослинними, тваринними і мінеральними царствами, з одного боку, людиною, її побутом і навіть духовним світом – з іншого. Саме ці співвідношення, ці закономірні взаємодії і складають суть пізнання єства (естества), ядро істиної натурфілософії (Докучаев, 1949, с. 317).
Початок практичного здійснення цієї широкої програми стосовно наземних природних комплексів пов’язано з ім’ям Г.Ф.Морозова (1867-1920 рр.) – засновника вчення про ліс. В своїй знаменитій праці він підкреслював, що ліс і його територія повинні зливатися для нас в єдине ціле, в географічний індивідум або ландшафт.
Ліс це цілий гуртожиток (общежитие) не лише рослинних, але і тваринних форм, що існують під владою зовнішнього географічного середовища і у зв’язку з ним. Таке цілісне уявлення про ліс передбачає необхідність вивчати як власне сам ліс, як такий, так і різні сторони його життя з умовами, що їх породжують (Морозов, 1925, с.319-320, 322-323).
Схожі ідеї висловлював Р.І.Аболін (1914) на основі вивчення боліт. Він вважав, що поверхнева оболонка Землі, названа ним «епігенемою» (що майже відповідає поняттю про екосферу чи біогеоценотичну оболонку) складається з окремих ділянок, які він називав «епіморфами» (приблизно відповідає поняттю про біогеоценози). В межах епіморфи рельєф, грунт, земля (почва) і рослинність поєднуються (сочетаются) і взаємодіють між собою, причому всередині її характер взаємодій зберігається відносно однорідним.
Енергію визначають як здатність виконувати роботу. Властивості енергії описуються двома законами термодинаміки. Перший закон термодинаміки, чи закон збереження енергії, проголошує, що енергія не зникає і не створюється заново, вона лише може переходити з однієї форми в іншу. Другий закон термодинаміки, або закон ентропії, формулюється по-різному, зокрема: процеси, пов’язані з перетворенням енергії, можуть протікати самоплинно лише за умови, що енергія переходить з більш концентрованої форми до більш розсіяної, тобто деградує. Другий закон можна сформулювати і так: будь-яке перетворення енергії з однієї форми у іншу неминуче супроводжується розсіюванням її частини у формі, недоступній для подальшого використання. Одним з наслідків цього є неможливість створення вічного двигуна.
Ентропія (від грецького entropia – перетворення, поворот?) – міра кількості зв’язаної енергії, яка стає недоступною для використання. Цей термін також використовується як міра зміни невпорядкованості системи, що відбувається при деградації енергії.
Енергетичний підхід до вивчення екологічних процесів значною мірою сприяв формуванню екології як науки, оскільки саме він дав змогу вивчати дивовижне розмаїття життя в усіх його проявах через встановлення кількісних закономірностей як міжорганізменних, так і міжпопуляційних взаємин, визначати енергетичні баланси біо- і екосистем різного рівня організації. Колосальна кількість публікацій цього напрямку сприяла з’ясуванню загальних закономірностей трансформації енергії як окремими організмами, популяціями, угруповуваннями, так і складових енергетичного балансу біосистем різного рівня організації. Але при цьому практично не враховується вплив якості середовища на жоден продукційно-енергетичний параметр. В практиці біопродукційних досліджень часто майже всі величини одержують розрахунковими методами, використовуючи продукційно-біомасові (Р/В) коефіцієнти, одержані (у більшості випадків) десятки років тому за умов зовсім іншого екотоксикологічного стану екосистем. Це не лише унеможливлює одержання об’єктивної інформації з продукційних параметрів різних видів, популяцій і трофічних рівнів, беручи до уваги значне погіршення стану середовища за останні десятиліття. Тим паче що за рівнем забруднення окремі екосистеми відрізняється настільки істотно, що його неврахування зводить нанівець цінність проведених таким чином біопродукційних досліджень.
|
|
|
|
|
Дата добавления: 2015-05-24; Просмотров: 511; Нарушение авторских прав?; Мы поможем в написании вашей работы!