КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
ФІЗІОЛОГІЯ М’ЯЗІВ
|
|
|
|
Термін «скорочення» означає взаєзв'язок збудження в скелетних м'язах (виникнення та поширення ПД по мембрані волокна) та його скорочення, тобто актоміозинової взаємодії. В стані спокою взаємодії актинових та міозинових протофібрил немає тому, що активні центри актину заблоковані регуляторними білками тропоніном та тропоміозином. Знімають блокаду іони Са2+, концентрація яких в саркоплазмі в стані спокою низька 10-8 ммоль/л.
ПД, що виникають в постсинаптичній мембрані поширюються вздовж всієї довжини мембрани м'язового волокна, у тому числі і по мембрані Т-трубочок (це вгинання зовнішньої мембрани м'язових клітин всередину волокна).
Безпосередньо поруч з Т-трубочками розташовані цистерни саркоплазматичного ретикулуму (СПР - частина розташованими цистернами утворюють тріади. СПР крім цистерн мають поздовжні трубочки, в яких локалізуються кальцієві насоси - вони активно транспортують іони - Са2+ із саркоплазми в СПР і в його цистернах накопичується велика кількість іонів Са +. При русі ПД по мембрані Т-трубочок в мембрані цистерн СПР відкриваються кальцієві канали і іони Са по градієнту концентрації виходять з цистерн СПР у саркоплазму з підвищення концентрації іонів Са2+ в саркоплазмі мюцита з 10-8 до 10-5 ммоль/л, дифузія іонів Са2+ до протофібрил, взаємодія з регуляторним білком тропоніном, зміна третинної конформації тропоніну та тропомюзину, відкриття активних центрів актину, взаємодія головок міозину з активними центрами актину (м'язове скорочення).
В основі м'язового скорочення лежить ковзання актинових протофібрил відносно міозинових "теорія ковзання".
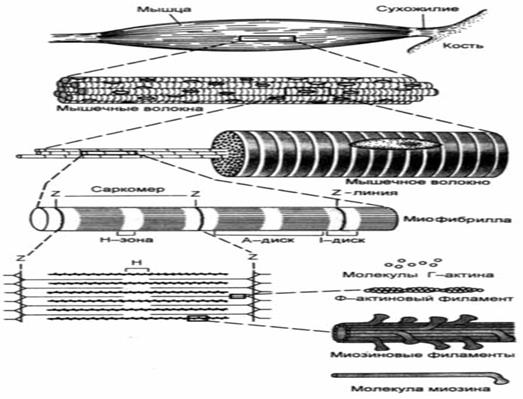
Саркомер – структурно -функціональний елемент скорочувального апарату скелетних м'язів. Вони містять актинові та міозинові нитки утворені пучками міофібрил, які відокремлені один від одного.
Саркомер в стані скорочення відокремлений перпендикулярними лініями смугами - Z-лініями. До Z-ліній прикріпляються одним своїм кінцем тонкі актинові нитки. Інші кінці
актинових ниток спрямовані до центру саркомера і входять у проміжки між товстими міозиновими нитками. Частина саркомера, яка примикає до Z-лінії
та утворена тільки актиновими протофібрилами, має назву І-дисків (ізотропних); слідом за ними розташовані А-диски (анізотропні) частина саркомера, де має місце взаємне перекриття актинових та міозинових протофібрил. При вкороченні м'яза, вході його скорочення, коротшає довжина протофібрил? вкорочення довжини кожного саркомера. Але при цьому довжина анізотропних дисків не зменшується, а зменшується довжина ізотропних дисків. Це є наслідком ковзання актинових протофібрил відносно міозинових в напрямку до центру саркомера.
Причиною руху акти нових ниток відносно міозинових (їх ковзання) є "нахил" голівки після її приєднання до активного центру актину.
Велика кількість голівок, що входять до складу актоміозинових містків, одночасно роблять "нахил" л нитка актину переміщується до центру саркомера вздовж нитки міозину. За рахунок одного такого "нахилу" саркомер (м'яз) може вкоротитися на 1% своєї вихідної довжини. Ступінь вкорочення м'яза може досягати 50% л такі "нахили" голівок під час одного м'язового скорочення мають повторитися 50 разів. Це можливо, якщо актоміозинові містки після "нахилу" розпадаються (дисоціюють) л голівка міозину взаємодіє з наступним активним центром актину л наступний "нахил" л розпад містку і т.д. На "нахил" однієї головки міозину витрачається енергія однієї молекули АТФ. Для того, щоб відбувся розпад актоміозинового містка необхідно, щоб до голівки міозину приєдналася молекула АТФ (але енергія АТФ при цьому не витрачається).
Для розслаблення скелетного м'язу необхідно, щоб зменшилась концентрація іонів Са2+ в саркоплазмі. Це досягається завдяки дії кальцієвих насосів повздовжніх трубочок СПР - активний транспорт іонів Са2+ в СПР – зменшення його концентрації в саркоплазмі до рівня 10-8 ммоль/л - від'єднання іонів Са від регуляторного білка тропоніну - відновлення вихідної третинної структури тропоніну та тропоміозину - блокада активних центрів актину - розслаблення м'язу.
Таким чином, під час м'язового скорочення енергія АТФ витрачається:
1. На роботу натрій-калієвого насосу зовнішньої мембрани м'язового волокна (цей насос підтримуєградієнт концентрації іонів К+ та Nа поза та всередині
клітини, що необхідно для підтримки на необхідному рівні ПС та для розвитку ПД на мембрані).
2. На "нахил" головки міозину.
3. На роботу кальцієвого насосу СПР - розслаблення м'язу.
11. Види м'язових скорочень: одинокі та тетанічні; і иноііічні ти ізометричні.
В залежності від режимів навантаження виділяють Наступні види м'язового скорочення.
1. Ізометричного скорочення - скорочення при незмінній довжині м'яза. Довжина м'яза при скороченні залишається сталою, але змінюється її напруження (Н). В експерименті таке скорочення можна отримати, якщо ізольований м'яз закріпити з двох сторін та стимулювати електричним струмом. В умовах цілісного організму ізометричне скорочення буває, коли людина намагається, але не може підняти вантаж.
 2. Ізотонічне скорочення - скорочення при незмінному напруженні м'яза. Напруження м'яза при його скороченні залишається сталим, а довжина змінюється (А1). В
2. Ізотонічне скорочення - скорочення при незмінному напруженні м'яза. Напруження м'яза при його скороченні залишається сталим, а довжина змінюється (А1). В
експерименті таке скорочення можна якщо один кінець ізольованого м'яза закріпити, а інший залишити вільним та стимуліоваги. В умовах цілісного організму скорочення м'язів максимально наближується до ізотонічного, якщо відбувається згинання ненавантажепих кінцівок. 3. Змішане (ауксотонічне) скорочення - скорочення, при якому змінюється довжина та напруження м'яза. Такий тип м'язових скорочень найчастіше зустрічається в умовах цілісного організму.
В залежності від частоти стимуляції виділяють наступні види м'язового скорочення:
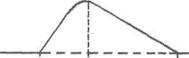
1- Одиночне - спостерігається у відповідь на одиничний стимул чи на їх серію (якщо інтервал між подразниками більший чи рівний тривалості одиночного м'язового скорочення). Виділяють такі періоди м'язового скорочення:
|
- латентий період (1) -
інтервал часу від дії подразника (П) до початку
скорочення;
1 2 3 - період скорочення (2) – під час цього періоду довжина м'яза зменшується;
- період розслаблення (3) - під час цього періоду м'яз розслаблюється, його довжина збільшується до вихідного рівня.
2. Тетанічне - спостерігається в умовах сумації одиночних м'язових скорочень (одне скорочення накладається на інше). Така сумація можлива, якщо м'яз подразнюється серією стимулів і кожен наступний стимул діє тоді, коли попереднє одиночне скорочення ще не закінчилось (тобто, інтервал між стимулами має бути меншим тривалості одиночного м'язового скорочення).
Виділяють наступні види тетанічних скорочень:
Зубчастий тетанус – виникає на базі одиночних скорочень, якщо кожен наступний стимул попадається на період розслаблення одиночного м’язового скорочення. Отже, інтервал між стимулами в серії має бути меншої тривалості одиночного м’язового скорочення, але більший, чим сума тривалості латентного періоду і періоду вкорочення одиночного скорочення.
Гладкий тетанус – виникає на базі одиночних скорочень, якщо кожен наступний стимул попадає на латентний період або період вкорочення одиночного м'язового скорочення. Отже, інтервал між стимулами в серії повинний бути меншим чи рівним сумі тривалості латентного періоду одиночного скорочення, але більше тривалості латентного періоду одиночного скорочення. Нижче зображений запис реєстрації скорочень м'яза при збільшенні частоти її стимуляції. Видно, що при збільшенні частоти стимуляції на фазі одиночних скорочень виникають тетанічні, спочатку зубчастий, а потім гладкий тетанус. Тетанічні скорочення відрізняється від одиночного наступними параметрами:
1. Тривалість тетанічного скорочення більша, ніж одиночного.
2. Амплітуда (сила) тетанічною скорочення більша, ніж одиночного. Амплітуда гладкого тетануса більша, ніж зубчастого.
Асинхронний тетанус. Асинхронний тетанус представляє вид скорочень, який притаманний скелетним м'язам при їх природній діяльності в умовах цілісного організму.
Кожен скелетний м'яз складається з великої кількості окремих м'язових волокон. При природній діяльності м'язів в умовах цілісного організму частота, з яким до них надходять ПД по аксонах мотонейронов недостатня для того, щоб у кожному окремому волокні м'яза розвивався гладкий тетанус. У той же час м'язи людини здатні до тривалого злитого скорочення. Таке скорочення є наслідком асинхронного (неодночасного) скорочення окремих м'язових волокон м'яза. Кожне волокно скорочується в режимі одиночних скорочень, але в межах м'яза відбувається (додавання) скорочень окремих волокон. У підсумку м'яз скорочується злитно та довготривало. Це і є асинхронний тетанус. Його тривалість може бути дуже великою, а його амплітуда не буває такою значною, як амплітуда гладкого тетанусу, який можна одержати в експерименті.
| Одиночні Зубчатий Гладкий скорочення тетанус тетанус |

|
Фізіологічні особливості гладких м'язів. Гладкі м'язи утворюють стінки (м'язовий шар) внутрішніх органів і кровоносних судин.
Мікроскопічна будова гладких і поперечносмугастих м'язів різна. У міофибрилах гладких м'язів немає смугастість. Це обумовлено хаотичним розташуванням скорочувальних білків у волокна гладких м'язів. Волокна гладких м'язів щодо коротше - від 50 до 200 мкм.
Фізіологічні властивості гладких м'язів у зв'язку з особливостями їх будови і рівня обмінних процесів значно відрізняється від фізіологічних властивостей поперечносмугастих м'язів.
Гладкі м'язи менш збудливі, ніж поперечносмугасті. Порушення по ним поширюється з невеликою швидкістю -2-15 см / с. Порушення в гладких м'язах може передаватися з одного волокна на інше, на відміну від нервових волокон і волокон поперечносмугастих м'язів.
Скорочення гладкої мускулатури відбувається більш повільно і довго. Внаслідок тривалості скорочувального акта гладка м'яз навіть під впливом окремих подразників може переходити в стан тривалого скорочення, яке нагадує тетанус скелетних м'язів. Характерними для гладких м'язів є також тривалі тонічні скорочення.
Рефрактерний період в гладких м’язах більш тривалий, ніж у скелетних (до декількох секунд).
Важливою властивістю гладкою м'язи є її велика пластичність, тобто здатність зберігати додану розтяганням довжину без зміни напруги. Дана властивість має суттєве значення, оскільки деякі органи черевної порожнини (матка, сечовий міхур, жовчний міхур) іноді значно розтягуються.
Характерною особливістю гладких м'язів є їх здатність до автоматичної діяльності, яка забезпечується нервовими елементами, закладеними в стінках гладком'язових органів.
Адекватним подразником для гладких м'язів є їх швидке і сильне розтягування, що має велике значення для функціонування багатьох гладком'язових органів (сечовід, кишечник та інші порожнисті органи).
Особливістю гладких м'язів є також їх висока чутливість до деяких біологічно активним речовинам (ацетилхолін, адреналін, норадреналін, серотонін та інші).
Гладкі м'язи іннервуються симпатичними і парасимпатичними вегетативними нервами, які, як правило, мають протилежний вплив на їх функціональний стан.
|
|
|
|
|
Дата добавления: 2015-06-04; Просмотров: 3892; Нарушение авторских прав?; Мы поможем в написании вашей работы!