КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Сплайсинг РНК может регулироваться таким образом, что один и тот же ген направляет синтез различных
|
|
|
|
10-23
форм белка [39]
Первоначально сплайсинг РНК был открыт у вируса. Оказалось, что из одного первичного транскрипта у него образуется несколько молекул мРНК, и, следовательно, синтезируется несколько разных белков (см. разд. 9.4.12). Многие гены высших эукариот образуют различные белки именно с помощью альтернативного сплайсинга РНК. Если в нескольких участках транскрнпта существуют различные точки сплайсинга, то один и тот же ген может служить матрицей для десятков различных белков. Обычно, однако, возможности сплайсинга ограничены и с каждого транскрипта транслируется лишь несколько белков.
В некоторых случаях альтернативный сплайсинг имеет место вследствие «двусмысленности интрона»: стандартный механизм удаления интронов не может четко различить две или более альтернативные пары 5'- и З'-сайтов сплайсирования, и, таким образом, в разных ситуациях случайно реализуются разные варианты. Подобная конститутивная форма альтернативного сплайсинга, по-видимому, ответственна за образование различных аномальных мРНК мутантного гена (3-глобина у некоторых лиц, страдающих β-талассемией (см. рис. 9-86). Для других генов такая двусмысленность характерна и в норме, при этом во всех тканях, где ген экспрессируется, образуются кодируемые им разные версии одного и того же белка.
Во многих случаях регулируется именно альтернативный сплайсинг РНК, а не конститутивный. Отбор сайтов сплайсинга определяется клеткой. Следовательно, в разных клетках в соответствии с потребностями организма с одного и того же первичного транскрипта РНК могут транслироваться различные белки (или наборы белков). Таким образом, многие белки образуются в тканеспецифичных формах. Среди них компоненты 1) внеклеточного матрикса (фибронектин), 2) клеточного скелета (тропомиозин), 3) плазматической мембраны, 4) ядра (см, табл. 10-1)
и 5) внутриклеточных путей клеточной сигнализации (С-киназа и тирозин-протеинкиназа, кодируемые протоонкогеном src; рис. 10-51).
Как правило, замены экзонов, происходящие при альтернативном сплайсинге РНК, не приводят к появлению совершенно разных белков.
Вместо этого образуется серия белков, функции которых аналогичны. Их называют изоформамн. Изоформы белка модифицированы таким образом, чтобы они годились для конкретной ткани. Изменения в них могут определять, с какими другими белками будет взаимодействовать данная молекула, причем каталитические или структурные домены
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
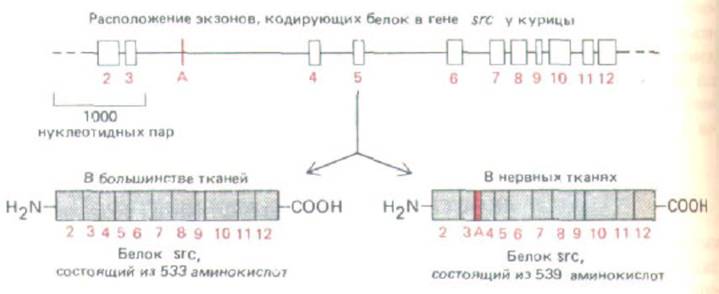
Рис. 10-51. При регуляторном альтернативном сплайсинге РНК с гена src считываются две несколько отличающиеся формы тирозин-
протеинкиназы. Экзон А включается в последовательность лишь в нервных тканях. Поскольку такая тканеспецифичность в сплайсинге сохранилась в ходе эволюции (характерна и для птиц, и для млекопитающих), можно предположить, что наблюдаемая разница в белках, кодируемых геном src, весьма важна для биологической активности этого регуляторного белка. На рисунке указаны только экзоны, кодирующие белок (экзон 1 формирует 5'-лидерную последовательность мРНК). (По J. В. Levy et al., Моl. Cell Biol. 7: 4142-4145, 1987.)
остаются прежними. Например, одна и та же транскрипционная единица в клетках щитовидной железы дает начало кальцитонину, а в клетках нервной ткани - далекому от него по свойствам пептидному гормон (CGRP - calcitonin-gene-related peptide).
10.4.3. Альтернативный сплайсинг РНК может использоваться для включения и выключения генов [40]
Некоторые гены постоянно транскрибируются во всех клетках, однако из-за существования конститутивного сплайсинга образуемая мРНК кодирует нефункционирующий белок, и ген экспрессируется лишь в тех клетках, где происходит специализированная реакция сплайсинга.
Такой тип генной регуляции особенно хорошо охарактеризовав у дрозофилы. Так, например, свойство Р-элемента (см. разд. 5.6.10)
перемещаться лишь в половых клетках, связано с его неспособностью образовывать активную транспозазу в соматических клетках. Это явление в свою очередь обусловлено присутствием в мРНК транспозазы
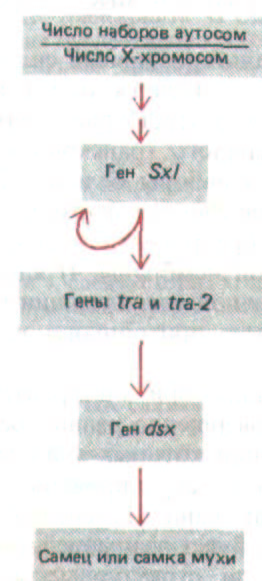
Рис. 10-52. Каскадные изменения в экспрессии генов, определяющих пол у дрозофилы, зависят от альтернативного сплайсинга РНК. Если отношение числа наборов аутосом к числу Х-хромосом равно единице (в норме это два набора аутосом и две Х-хромосомы), особь развивается в самку; мухи, у которых это отношение равно двум (в норме два набора аутосом и одна Х-хромосома), развиваются как самцы. Это соотношение определяется уже на ранних стадиях развития и затем сохраняется в каждой клетке, Функция генов, представленных на рисунке, как раз и состоит в передаче информации об этом соотношении другим генам, определяющим фенотип, характерный для того или иного пола. Такие гены функционируют как два альтернативных набора, определяющие свойства, характерные для самки и для самца. Ген dsx (doubl esex - двупол ость)
получил свое название благодаря тому, что у мутантов, не экспрессирующих этот ген, работают оба набора, специфичные для самок и для самцов.
На рисунке представлена цепь событий, которые происходят при развитии самок. Стрелки указывают действие каждого гена, участвующего в этом процессе. У самцов гены Sxl, tra и tra2 транскрибируются, но образуют только нефункциональную мРНК, а транскрипт гена dsx сплайсируется, причем в итоге образуется белок, который включает гены, детерминирующие женский путь развития. У самок транскрипт Sxl
сплайсируется совершенно по-другому, в результате образуется белок, контролирующий сплайсинг, который, с одной стороны, поддерживает свой собственный синтез, а с другой стороны, включает два гена tra (указано стрелками). В свою очередь, продукты генов tra, действуя кооперативно, меняют схему сплайсинга РНК у транскрипта гена dsx. Получившаяся мРНК гена dsx образует измененную форму белка dsx, которая выключает гены, определяющие мужской путь развития.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
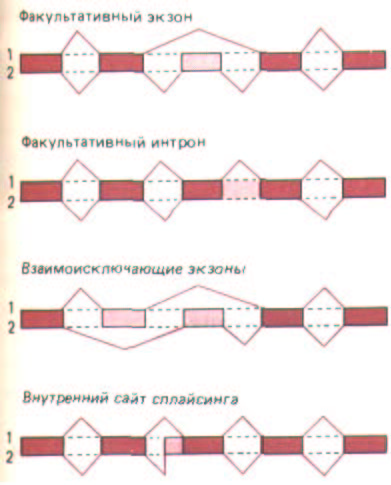
Рис. 10-53. Четыре пути альтернативного сплайсинга РНК, которые имели место в эксперименте. В каждом случае одна транскрибированная РНК может сплайсироваться двумя альтернативными способами с образованием двух разных мРНК (1 и 2), Более темным цветом обозначены последовательности РНК, присутствующие в обоих типах мРНК. Прямоугольники, окрашенные светлее, соответствуют последовательностям, имеющимся лишь в мРНК одного типа. Соседние прямоугольники соединены цветными линиями, указывающими на последовательности интронов. По-видимому, не существует какого-либо простого механизма или единого правила, которые могли бы объяснить, почему происходит тот или иной выбор. (По A. Andre-adis, М. Е. Gallego, В. Nadal-Ginard, Annu. Rev. Cell Biol. 3: 207-242, 1987.)
интрона, который, по-видимому, удаляется лишь в половых клетках. С помощью генетического анализа выявлен другой пример. Пол мух определяется каскадной активацией генов, каждый из которых ответствен за синтез белка, детерминирующего правильный сплайсинг РНК следующего в этом ряду гена (рис. 10-52). Области ДНК, кодирующие некоторые белки, которые детерминируют пол мухи, клонированы и секвенированы, что в значительной мере облегчило изучение механизмов, регулирующих выбор сайта сплайсинга.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 644; Нарушение авторских прав?; Мы поможем в написании вашей работы!