КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Присутствие энхансера трансляции в некоторых вирусных мРНК свидетельствует о существовании
|
|
|
|
механизма позитивного контроля трансляции [45]
В принципе, позитивный контроль трансляции может осуществляться засчет расположенной на мРНК специальной области «трансляционного энхансера», который способен выборочно привлекать рибосомы. Показано, что определенные РНК-вирусы (пикорнавирусы)
содержат такую область. Ее присутствие приводит к тому, что трансляция начинается с внутренних сайтов AUG, которые в других случаях в эукариотической клетке не используются для инициации синтеза белка (рис. 10-56).
Позитивный контроль трансляции обнаружен также и в дрожжевых клетках. Генетическими методами были выявлены специфические белки, необходимые для активации трансляции мРНК дрожжевого гена GCN4. В отсутствие этих белков мРНК не транслируется. мРНК гена GCN4
напоминает слабо транслируемые мРНК высших эукариот. Полагают, что трансляция таких мРНК контролируется аналогичным образом. (К этому классу относится около 5% всех охарактеризованных до сих пор мРНК). У таких РНК 5'-лидерная последовательность необычно длинна и содержит серию триплетов AUG, препятствующих трансляции основной кодирующей последовательности, расположенной за сайтом инициации синтеза коротких пептидов. Стоп-кодон, локализованный перед основной кодирующей последовательностью, препятствует сквозному считыванию.
Сходным образом, у пикорнавирусов трансляция основной
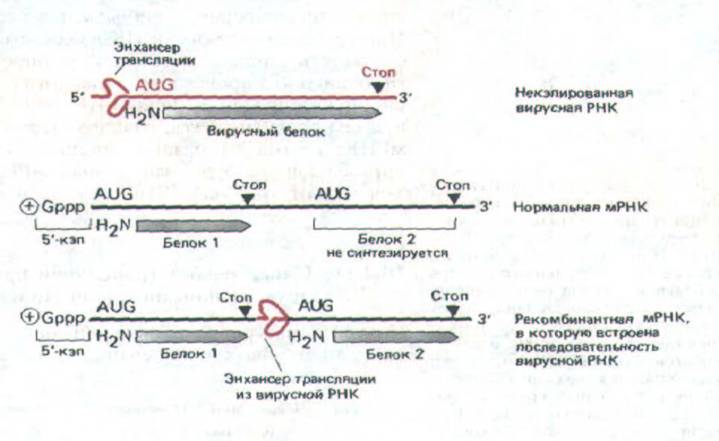
Рис. 10-56. Схема эксперимента, который демонстрирует существование последовательности РНК в геномах некоторых РНК-содержащих вирусов, выступающей в роли трансляционного энхансера. Геном пикорнавирусов (к которым относится и вирус полиомиелита) представляет собой плюс-
нить РНК, т.е. может служить непосредственно мРНК для синтеза вирусоспецифических белков. У этих вирусов нет нэпа на 5'-конце РНК, который необходим для инициации белкового синтеза на большинстве молекул мРНК. Измеряя уровень белкового синтеза, катализируемого различными рекомбинантными РНК, в молекуле вирусной РНК можно идентифицировать последовательности трансляционных энхансеров длиной несколько сот нуклеотидных пар. Как показано на схеме, перемещая эту последовательность в середину цепи мРНК, можно вставить рибосому начинать трансляцию с соседнего внутреннего кодона AUG; таким образом, можно обойти правила, которые в норме заставляют начинать белковый синтез только с первого кодона AUG.
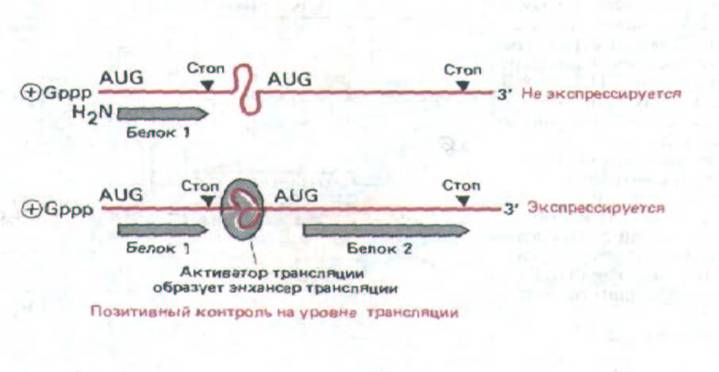
Рис. 10-57. Модель позитивного контроля трансляции, согласно которой для интенсивной трансляции мРНК необходимо связывание белка (активатора трансляции). Хотя известно, что трансляция специфических мРНК находится под позитивным контролем, его механизм до конца неясен. Установлено, что позитивный контроль имеет отношение к синтезу коротких пептидов, трансляция которых инициируется перед первым AUG. Это дает право считать, что в данном случае действует механизм, аналогичный тому, который описан у пикорнавирусов (рис. 10-56).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
кодирующей последовательности такой мРНК может зависеть от связывания молекул-активаторов трансляции с последовательностями энхансеров трансляции, расположенных вблизи соответствующих кодо-нов AUG, что приводит к повторной инициации трансляции (рис. 10-57). Однако каков механизм подобной активации трансляции, до сих пор неизвестно.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 418; Нарушение авторских прав?; Мы поможем в написании вашей работы!