КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Изменение сайта, в котором происходит расщепление транскрипта РНК и его полиаденилирование, может
|
|
|
|
Механизмы, ответственные за выбор сайта для регулируемого сплайсинга РНК, неизвестны
10-23
Полагают, что регулируемые изменения в выборе сайтов сплайсинга РНК осуществляются путем связывания ткане- и геноспецифичных белков или молекул РНК с растущим РНК-транскриптом. Так как выбор сайтов сплайсинга происходит и при конститутивном, и при регулируемом варианте по одним и тем же стандартным консенсусным последовательностям, связывание с определенным компонентом должно менять конформацию транскрипта РНК, с тем чтобы закрыть или открыть сайты сплайсинга, ранее существовавшие в молекуле. Вероятно, тут задействованы сложные механизмы, так как простое предположение о том, что при связывании белка сайт сплайсинга закрывается, не объясняет в достаточной мере, почему наблюдается такое разнообразие продуктов сплайсинга (рис. 10-53). Для изучения молекулярных механизмов регулируемого сплайсинга необходимо реконструировать его в бесклеточной системе, что позволило бы выделить все необходимые компоненты и проанализировать действие каждого из них на сплайсосому.
менять карбоксильный конец белка [41]
У эукариот З'-конец молекулы мРНК определяется не терминацией синтеза РНК РНК-полимеразой, а в ходе реакции расщепления РНК, которая катализируется дополнительными факторами при элонгации транскрипта (см. разд. 9.4.5). Место такого расщепления может варьировать, при этом изменяется карбоксильный конец получающейся молекулы белка, который кодируется З'-концом мРНК. У прокариот образование более длинного транскрипта РНК приводит лишь к появлению дополнительных аминокислот в белковой цепи. Однако у эукариот при сплайсинге образование более длинного транскрипта может привести к тому, что исходный карбоксильный конец белка будет полностью удален и замещен новым.
С изменением такого типа связано переключение синтеза антител при созревании В-лимфоцитов с мембраносвяз энных на секретирующие формы. В незрелых В-клетках образующиеся антитела связаны с плазматической мембраной, где они служат рецепторами антигенов. Стимуляция антигенами одновременно запускает деление этих клеток и начало секреции ими антител. Секретируемая форма антител отличается от мембраносвязанной только в терминальной части карбоксильного конца: мембраносвязанная форма содержит здесь длинную цепь из гидрофобных аминокислот, пересекающую липидный бислой, а секретируемая форма несет гораздо более короткий фрагмент водорастворимых аминокислот. Таким образом, для переключения с синтеза мембраносвязанных на секретируемые антитела необходима иная нуклеотидная последовательность на З'-конце мРНК.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
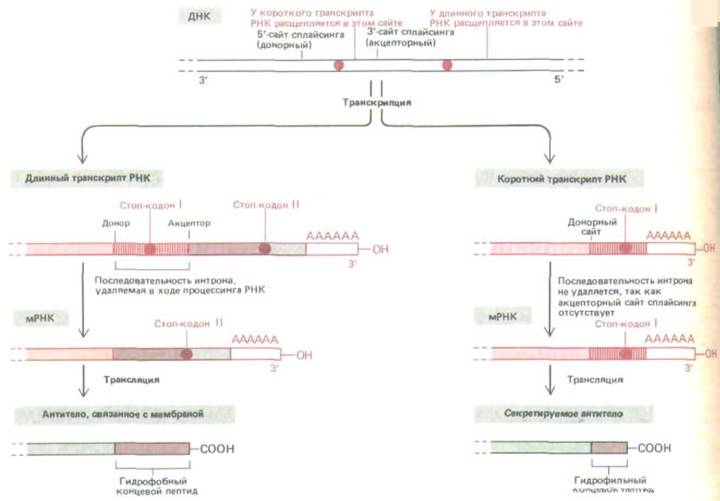
Рис. 10-54. Выбор сайта расщепления РНК и полиаденилирования весьма важен при образовании антител. В нестимулированных В-
клетках (слева) образуются длинные транскрипты РНК, и при сплайсинге удаляется последовательность интрона вблизи З'-конца этой молекулы. В результате образуется мРНК, которая кодирует молекулу антитела, связывающуюся с мембраной. Напротив, после стимуляции антигеном (справа)
первичный транскрипт РНК разрезается перед акцепторным сайтом сплайсинга последнего экзона. В результате некоторые последовательности интрона, удалявшиеся из длинного транскрипта, остаются в качестве кодирующих последовательностей в коротком транскрипте. Это те последовательности, которые кодируют гидрофильную карбоксиконцевую часть секретируемой молекулы антитела.
Мембраносвязанная форма белка образуется при транскрипции всех кодирующих последовательностей ДНК с образованием длинного транскрипта. Нуклеотиды, кодирующие длинный гидрофобный карбоксильный конец мембраносвязанного белка, локализованы в последнем экзоне (рис. 10-54, слева). Интрон, предшествующий этому экзону, содержит нуклеотиды, кодирующие водорастворимый хвост секретируемой молекулы;
при сплайсинге мРНК они удаляются. Секретируемая форма молекулы образуется с более короткого первичного транскрипта, который заканчивается перед началом следующего экзона. Следовательно, в этом транскрипте перед нуклеотидами, кодирующими водорастворимый хвост, отсутствует акцепторный сайт сплайсинга, который мог бы взаимодействовать с имеющимся донорным сайтом. В результате эти нуклеотиды остаются в образующейся молекуле мРНК (рис. 10-54, справа).
Неизвестно, однако, как контролируется реакция расщепления, определяющая переключение длины транскриптов РНК.
10.4.6. Открытие альтернативного сплайсинга требует пересмотра понятия «ген» [42]
С тех пор, как стало известно, что эукариотические гены содержат интроны, а их кодирующие последовательности можно состыковать по-разному, вновь встал вопрос о том, что следует понимать под словом «ген». Первое определение гена на молекулярном уровне было предложено в начале 40-х гг. на основании изучения биохимической генетики гриба
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
нейроспора. До этого времени геном считали область генома, которая в мейозе обособляется как отдельная единица и ответственна за проявление определенного фенотипического признака, например, белых и красных глаз у дрозофилы или гладких и морщинистых семян у гороха. После работ на нейроспоре стало ясно, что ген, как правило, соответствует области генома, направляющей синтез единственного фермента. На основании этого возникло предположение, что один ген кодирует одну полипептидную цепь. Эта гипотеза оказалась весьма плодотворной. При изучении механизмов экспрессии в 60-х гг. ген стали определять как фрагмент ДНК, который транскрибируется с образованием РНК, кодирующей одну полипептидную цепь (или одну структурную РНК, как например, молекулы тРНК или рРНК). Обнаруженный в конце 70-х гг. феномен прерывистости генов эукариот не противоречил принятым представлениям. Однако теперь нам стало ясно, что в клетках высших эукариот многие последовательности ДНК могут кодировать два и более различных белка благодаря альтернативному сплайсингу. Что же тогда следует считать геном?
В тех относительно редких случаях, когда два сильно различающихся эукариотических белка образуются из одной транскрипционной единицы, говорят, что эти белки кодируются различными генами, которые на хромосоме перекрываются. Однако определение видоизмененных белков, образующихся в результате альтернативного сплайсинга РНК, как продуктов перекрывающихся генов, может показаться излишне усложненным. Проще изменить исходную формулировку и считать геном любую последовательность ДНК, которая транскрибируется как отдельная единица и кодирует набор близкородственных полипептидных цепей (изоформы белков).
10.4.7. Экспорт РНК из ядра может регулироваться [43]
Первичный транскрипт РНК в среднем в 10 раз длиннее, чем зрелая молекула РНК, образующаяся при сплайсинге. Но по некоторым данным пределы клеточного ядра покидает только одна двадцатая часть всей гяРНК (см. разд. 9.4.8). Таким образом, значительная часть первичных транскриптов (вероятно, половина) полностью разрушается в ядре, никогда не образуя молекулу мРНК, предназначенную для переноса. Распаду подвергаются и те молекулы РНК, последовательности которых не могут превратиться в мРНК, и такие молекулы, которые способны соответствующим образом процессироваться лишь в клетках другого типа.
Перенос РНК через ядерные поры представляет собой активный процесс (см. разд. 8.3.3). Если этот процесс зависит от специфического узнавания транспортируемых молекул РНК (либо связавшихся с РНК молекул белка или РНК) белком-рецептором, входящим в состав комплекса ядерной поры, то РНК, не имеющие такого опознавательного знака, будут избирательно задерживаться в ядре. С другой стороны, для переноса РНК могут и не потребоваться опознавательные сигналы, в этом случае автоматически переносится вся РНК, за исключением той, которая избирательно задерживается. Третья возможность заключается в том, что используется и селективный перенос, и селективное задержание. Поскольку любая РНК задерживается в ядре до тех пор, пока с ней связаны компоненты сплайсосомы, селективное задержание может обусловливаться механизмом, препятствующим завершению сплайсинга определенной молекулы РНК. В настоящее время ни одна из этих гипотез не получила четкого экспериментального подтверждения, более того, представляется маловероятным, чтобы транспорт РНК из ядра
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
играл существенную роль в регуляции экспрессии большинства генов у эукариот.
В связи с тем, что вирус использует для своего размножения клеточный аппарат, изучение цикла развития вируса часто помогает понять механизмы клеточных процессов. Например, геном аденовируса представляет собой двухцепочечную молекулу ДНК, которая реплицируется и транскрибируется в ядре клетки-хозяина. На поздних стадиях инфекции транспорт хозяйской РНК из ядра останавливается, в результате большая часть РНК, попадающей в цитоплазму, оказывается принадлежащей аденовирусу. Генетический анализ показал, что для такого изменения в избирательности переноса РНК из ядра необходимы два образующихся на ранних стадиях инфекции аденовирусных белка. Таким образом, взаимодействие аденовируса с клеткой-хозяином может служить перспективной модельной системой для изучения контроля за транспортом РНК.
10-24 10.4.8. Белки, связывающиеся с 5'-лидерной областью мРНК, участвуют в негативном контроле трансляции
[44]
Не все молекулы РНК, достигающие цитоплазмы, транслируются в белок. Трансляция некоторых из них блокируется особыми белками-
репрессорами (рис. 10-55), которые связываются вблизи 5'-конца, там, где должна происходить инициация трансляции. Такой тип регуляции был впервые обнаружен у бактерий, где избыток рибосомных белков может подавлять трансляцию своих собственных мРНК.
В клетках эукариот особенно хорошо изучен тип негативного контроля трансляции, который дает возможность быстро привести в соответствие синтез внутриклеточного белка ферритина с количеством имеющихся в клеточном содержимом атомов железа. Показано, что после добавления железа мРНК ферритина в цитоплазме перемещается из неактивного рибонуклеопротеинового комплекса в трансляционноактивный полирибосомный комплекс. С помощью генной инженерии установлено, что регуляция, направляемая железом, связана с последовательностью размером 30 нуклеотидов, которая расположена на 5'-лидерном конце молекулы мРНК ферритина. Этот элемент, контролирующий ответ на железо, складывается в структуру, состоящую из стебля с петлей (см. рис. 10-60, Б); элемент связывается с регуляторным белком в том случае, если этот белок не соединен с железом. При связывании регуляторного белка с элементом, контролирующим ответ на железо, трансляция любой последовательности РНК, расположенной за этим сайтом, подавляется (рис. 10-55). Добавление железа приводит к диссоциации комплекса РНК—
белок, что вызывает увеличение скорости трансляции мРНК в 100 раз.
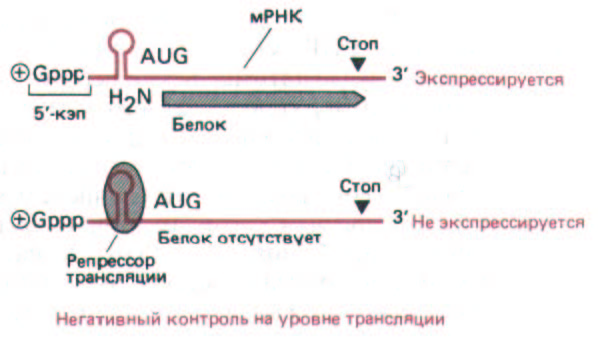
Рис. 10-55. Негативный контроль трансляции с участием сайт-специфического ДНК-связывающего белка (репрессор трансляции).
Связывание этого белка с молекулой мРНК приводит к снижению уровня ее трансляции. Известно несколько примеров контроля на уровне трансляции такого типа. В данном случае представлен механизм повышения синтеза ферритина при увеличении концентрации свободного железа в клетке (см. также рис. 10-60).
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 555; Нарушение авторских прав?; Мы поможем в написании вашей работы!